Реакция - фотосинтез
Cтраница 2
Более заманчивый способ восполнения дефицита энергии при реакции фотосинтеза заключается в использовании параллельно идущей реакции окисления кислородом. [16]
ТДФ в составе транскетолазы является одним из катализаторов реакций фотосинтеза у растений. Синтезируемый в пентозном цикле D-рибозо-б - фос-фат ( CLXXXI) является источником D-рибозы в реакциях биосинтеза рибонуклеиновых кислот ( РНК), АТФ, НАД, ФАД и других соединений. [17]
Так как даже в наиболее продуктивных сообществах в реакциях фотосинтеза используется всего 1 - 2 % солнечной энергии, то они не дают достаточно продукции, чтобы прокормить растущее человечество. Обратные соотношения - относительно малая биомасса и высокая первичная продуктивность - свойственны аг-робиоценозам, которые являются экономически выгодными. Однако без постоянного ухода и зашиты со стороны человека они быстро сменяются малопродуктивными природными биогеоценозами. [18]
Один из путей выяснения природы участия хлорофилла в реакциях фотосинтеза состоит в изучении кинетики и механизма простых ( но сравнению со всем процессом фотосинтеза) сенсибилизированных реакций, в которых участвует хлорофилл. [19]
В чем заключается основное отличие между световой и темновой реакциями фотосинтеза. [20]
За счет энергии Солнца происходит построение органического вещества в реакциях фотосинтеза ( хемосинтеза), т.е. энергия Солнца накапливается в веществе - экосистеме. [21]
Со времен Энгельмана и Рейнке стало общепринятым, что последовательность реакций фотосинтеза начинается и кончается в хлоро-пластах, несмотря на случайные, довольно смутные рассуждения о протоплазматическом факторе как регуляторе фотосинтеза. Однако неспособность отдельных, хотя бы и неразрушенных на вид, хлоро пластов осуществлять фотосинтез ( см. главу IV) показывает, что этот процесс требует соучастия цитонлазмы. [22]
На основании этих данных Кальвин делает следующие заключения о природе реакций фотосинтеза. [23]
В хлоропластах, присутствующих только Б растительных клетках, протекают все реакции фотосинтеза, как световые, так и темновые. При помощи хлоропластов клетки зеленых растений улавливают энергию солнечного света я превращают ее в химическую энергию. Каждый хлоропласт окружен оболочкой из двойной мембраны и содержит сложную внутреннюю систему мембран, разделяющую В. [24]
Ее тепловой эффект равен по величине и противоположен по знаку эффекту реакции фотосинтеза. [25]
Для объяснения образования дисилокеана и азота, а также повышенной скорости образования водорода в реакции фотосинтеза метилсиланов CHsSiHa, ( CHabSif - b, ( СН3) з5Ш в присутствии окиси азота высказано. [26]
Анализ световых кривых показал, что фосфаты повышают интенсивность как фотохимических, так и темно-вых реакций фотосинтеза. [27]
В результате фотосинтеза из 5 молекул образуется 6 молекул фосфоглицериновой кислоты, иначе говоря, итог реакций фотосинтеза заключается в превращении 3 молекул С02 в 1 молекулу фосфоглицериновой кислоты. [28]
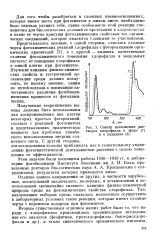 |
Спектр поглощения творов хлорофилла в и в пиридине. [29] |
Получаемые теоретические выводы должны быть использованы для воспроизведения вне клетки некоторых простых фотореакций, сходных с реакцией фотосинтеза и представляющих практическую ценность для проблемы химического использования энергии солнечного света. [30]
Страницы: 1 2 3 4