Значение - радиус - инерция
Cтраница 1
Значения радиуса инерции для частиц различной формы были приведены в разд. [1]
Значения радиусов инерции различных стандартных профилей прокатных балок приведены в приложении. [2]
В табл. 11 даны значения радиусов инерции для наиболее часто встречающихся типов сечения. [3]
При проектировании элементов фермы в расчет вводят значения радиусов инерции относительно свободной оси сечения. В зависимости от принимаемого типа сечения, например по рис. 3.29, б или по рис. 3.29, в, оси могут менять свои обозначения. Обычно принимают обозначение горизонтальной ( материальной) оси через х и вертикальной ( свободной) через у. [4]
В справочниках по моментам инерции приводят таблицы значений радиусов инерции различных тел. [5]
Для определения величины 1г воспользуемся тем обстоятельством, что для порожних грузовых автомобилей одной весовой категории и одного компоновочного типа значения радиусов инерции подрессоренной массы приблизительно одинаковы. [6]
Там Же даны значения радиусов инерции для составных элементов. [7]
Элементы замкнутых сечений обладают большей местной устойчивостью по сравнению с открытыми. В табл. 18 приведены значения радиусов инерции некоторых типов сечений стоек. [8]
Колеса, блоки, катки, для которых значения радиусов инерции в табл. 9 не указаны, считаются сплошными однородными дисками. [9]
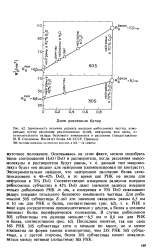
Экспериментально найдено, что нейтронное рассеяние белка скомпенсировано в 40 42 % DsO, в то время как РНК не видна для нейтронов в 70 % DzO. Соответственно измерения радиусов инерции рибосомных субчастиц в 42 % DsO дают значения радиуса инерции только рибосомной РНК in situ, а измерения в 70 % DsO показывают радиус инерции тотального белкового компонента частицы. РНК в виде ядра сосредоточена преимущественно у центра, а белок в среднем занимает более периферическое положение. Последнее понятно, так как сама 16S РНК 30S субчастицы хотя и меньше по массе, но и менее компактна по форме ( менее изометрична), чем 23S РНК 50S субчас-тицы, а с другой стороны, белок 30S субчастицы может локализоваться между ветвями ( лопастями) 16S РНК. [11]
Введение представления о существовании специфических изомерных состояний полимерной цепочки ( нем. Пеххольда не только учитывает упорядоченность аморфного состояния полимеров, но также, судя по результатам модельных расчетов [102], позволяет в принципе достичь количественного совпадения между значениями радиуса инерции меандровой структуры и невозмущенного статистического клубка, однако, как показали измерения интенсивности рассеяния деполяризованного света стеклообразными полиметилметакрилатом [105], поликарбонатом и полистиролом [106], максимальный размер обнаруженных анизотропных структур ( порядка 28 А [105]), а также степень ориентационнои корреляции макромолекул существенно меньше [105, 106], чем предсказывается меандровой моделью. [12]
Изложенные данные позволяют сделать вывод о негауссовом рлспределении звеньев макромолекулы, находящейся в конформации невозмущенного клубка. Этот вывод полностью согласуется с результатами исследования конформационного состояния макромолекул линейного полиэтилена в твердом состоянии методом малоуглового рассеяния нейтронов, которые свидетельствуют об идентичности как формы кривой рассеяния, так и значений радиуса инерции дейтерированных макромолекул в расплаве и кристаллическом состоянии. Поскольку тщательно продуманная методика приготовления образцов, по-видимому, исключает влияние возможных экспериментальных артефактов типа образования кластеров дейтерированных цепей, можно сделать заключение, что при кристаллизации в условиях быстрого охлаждения макромолекула в основном сохраняет ту же конформацию, что и в расплаве. [13]
Это расстояние рг У JZ / M называют радиусом инерции тела относительно оси. Радиус инерции измеряют в единицах длины. Выражение момента инерции тела через радиус инерции удобно потому, что это позволяет по значению радиуса инерции, указанному в таблице из справочников, найти момент инерции, учтя материал тела. [14]
Страницы: 1