Регуляция
Cтраница 1
Регуляция на уровне трансляции у прокариот практически целиком сводится к разрешению или предотвращению инициации трансляции данных кодирующих последовательностей мРНК рибосомами; именно этим путем достигается избирательная или преимущественная трансляция одних мРНК или отдельных ее цистронов и выключение трансляции других. Кроме того, разная скорость инициации на разных мРНК или их цистронах определяет соотношение продукции соответствующих белков. [1]
Регуляция может осуществляться на многих уровнях, но главную роль играют регуляторные механизмы двух типов. Один из них основан на том, что состав окружающей среды влияет на скорость и интенсивность синтеза различных ферментов. Этот механизм, относительно медленно действующий, регулирующий обмен путем индукции и репрессии, описан в гл. Следует обратить внимание, что скорости синтеза и распада регуляторных ферментов чаще всего регулируются гормонами. [2]
Регуляция идет путем воздействия на ключевой фермент биосинтеза конечным продуктом рибофлавином, ФМН, ФАД. [3]
Регуляции на всех уровнях связаны с предвосхищением. Вышестоящим, определяющим1 предвосхищение является предвосхищение общественно необходимого продукта трудовой деятельности. [4]
Регуляция осуществляется на многих уровнях, начиная с экспрессии генов. В одних клетках тот или иной канальный белок синтезируется, в других-нет. После синтеза этот белок направляется к надлежащему участку плазматической мембраны. Как н другие мембранные белки, он подвергается затем обновлению путем расщепления и замены вновь синтезированными молекулами. Скорость этого обновления весьма изменчива и может специально регулироваться. Обычно канальный белок сохраняется в мембране несколько дней, прежде чем перейдет в цитоплазму путем эндоцитоза и будет расщеп лен лн-зосомными ферментами. За гораздо более короткие промежутки времени, измеряемые уже минутами, а не часами и днями, изменяется число многих функционально активных каналов в результате ковалентной модификации их молекул: канальные белки могут фосфорнлироваться или метилироваться, что приводит к открытию или закрытию каналов. [5]
Регуляции) осуществляется взаимодействие между частями зародыша, необходимое для детерминации зачатка ЦНС. [6]
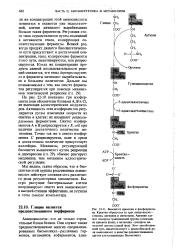
Регуляция этого типа осуществляется путем изменений в активности генов, кодирующих соответствующие ферменты. Всякий раз, когда продукт данного биосинтетического пути присутствует в достаточной концентрации, гены, кодирующие ферменты этого пути, инактивируются, или репрессируются. Когда же концентрация продукта данной последовательности реакций снижается, эти гены дерепрессируют-ся и ферменты начинают вырабатываться в большем количестве. Дальше мы познакомимся с тем, как синтез ферментов регулируется при помощи механизмов генетической репрессии ( гл. [8]
Регуляция на уровне активности ферментов свойственна, как правило, только ключевым ферментам клеточного метаболизма. При ингибировании конечным продуктом ( ре-троингибировании) этот продукт подавляет активность первого фермента, участвующего в данной цепи реакций. [9]
Регуляция многих физиолого-биохимических процессов в растениях совершается при участии комплекса эндогенных регуляторов роста, состав которых существенно изменяется в онтогенезе, в соответствии с чем меняется характер и направленность обмена веществ. Поэтому вполне оправдан интерес исследователей к познанию эндогенных регуляторов роста и изучению их действия на растения. [10]
Регуляция ионами Са2 осуществляется прямой активацией киназы фосфорилазы Ь без начального этапа, протекающего с участием протеинкиназы. [11]
Регуляция носит сложный характер. Фруктозе-1 6-дифосфат, образующийся под действием фосфофруктокиназы, активирует пируваткиназу по типу предваряющего эффекта. АТФ действует как мощный ингибитор по типу обратной связи ( стр. [12]
Регуляция этого типа характерна для анаболических систем, в особенности для тех, продукты которых представляют собой субстраты для синтеза наиболее важных биополимеров - белков и нуклеиновых кислот. Угнетающим агентом, концентрация которого контролирует скорость действия системы, является ее конечный продукт, промежуточные продукты таким действием не обладают. Эт избирательность обеспечивает целенаправленность регуляции. Конечный продукт не влияет ва активность. [13]
Регуляция гидра-туры ( показателем которой служит осмотическое давление клеточного сока) может достигаться либо приспособлениями корневой системы, обеспечивающими постоянный приток воды, либо путем ограничения транспирации. Определенное значение имеет и запасание воды в клетках и даже на уровне целого организма. [14]
Регуляция их на протяжении годового цикла важна не только в плане приуроченности каждого процесса к экологически оправданному сезону, но и в плане упорядоченного распределения физиологических состояний во времени. Это важно, так как совмещение энергоемких процессов биологически невыгодно. [15]
Страницы: 1 2 3 4