Регуляция - дыхание
Cтраница 2
Применение более сложных тестов и функциональных нагрузок позволило установить, что отравление сероводородом отражается на регуляции дыхания и-сердечной деятельности. Ухудшение регуляции дыхания, кратковременное и компенсирующееся, после вращения в центрифуге было найдено у крыс, отравленных меньшей концентрацией сероводорода. [16]
Регуляция дыхания обеспечивает две группы процессов. [17]
Исследовалась регуляция дыхания и кровообращения при функциональных нагрузках ( ортостатическая проба, вращение в центрифуге), определялась активность дегидраз в тканях методом Тунбер-га, применялись такие интегральные тесты, как частичное голодание на фоне отравления, плавательная проба, измерялось кровяное давление, регистрировалась ЭКГ, проба Торна и велись наблюдения за общим состоянием и весом животных. [18]
В системах с прочным сопряжением дыхание зависит от присутствия фосфата и АДФ. Такая регуляция дыхания обнаружена почти так же давно, как и фосфорилирование в дыхательной цепи. [19]
Применение более сложных тестов и функциональных нагрузок позволило установить, что отравление сероводородом отражается на регуляции дыхания и сердечной деятельности. Ухудшение регуляции дыхания, кратковременное и компенсирующееся, после вращения в центрифуге было найдено у крыс, отравленных меньшей концентрацией сероводорода. [20]
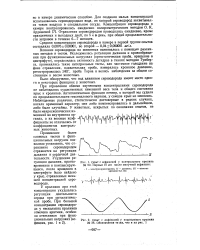 |
Опыт с асфиксией у контрольного кролика № 20. Первые 10 сек яосле минутной асфиксии. [21] |
Применение более сложных тестов и функциональных нагрузок позволило установить, что отравление сероводородом отражается на регуляции дыхания и-сердечной деятельности. Ухудшение регуляции дыхания, кратковременное и компенсирующееся, после вращения в центрифуге было найдено у крыс, отравленных меньшей концентрацией сероводорода. [22]
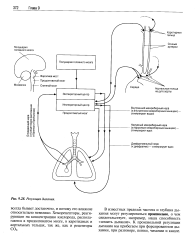 |
Регуляция дыхания. [23] |
В известных пределах частота и глубина дыхания могут регулироваться произвольно, о чем свидетельствует, например, наша способность затаить дыхание. К произвольной регуляции дыхания мы прибегаем при форсированном дыхании, при разговоре, пении, чихании и кашле. [24]
Здесь рассматривается только частный вопрос о влиянии аэроионов на дыхательную функцию и процессы газообмена, с одной стороны, и на окислительно-восстановительные явления и электрические свойства крови и тканей - с другой. Известно, что в регуляции дыхания существеннейшее значение имеют различные рефлексы, в основном рефлексы, вызываемые раздражением интерорецепторов. Изменение в функции дыхания, интенсивность газообмена и окислительно-восстановительные явления находятся в определенной зависимости от полярности и числа аэроио-нов во вдыхаемом воздухе и потому представляют на данном этапе разработки вопроса исключительный интерес. [25]
Центры гипоталамуса играют большую роль в регуляции дыхания во время поведенческих актов. Вследствие влияния центров гипоталамуса на дыхательный центр происходит усиление дыхания при общей защитной реакции организма, в частности при болевых раздражениях, во время физической работы, при эмоциональном возбуждении. [26]
В жизнедеятельности высших животных организмов основное значение имеют механизмы нервной и гормональной регуляции, преимущественно также на основе принципа О. Этим путем обеспечиваются го-меостатич. При регуляции дыхания повышение содержания С02 в альвеолах легких усиливает возбуждение нервных центров дыхания и интенсивность газообмена, что приводит к снижению содержания С02 в легких до нормы. Регулирование мозжечком непроизвольных мышечных движений происходит по принципу О. [27]
Хеморецепторы, стимулируемые увеличением напряжения двуокиси углерода и снижением напряжения кислорода, находятся в каротидных синусах и дуге аорты. Они расположены в специальных маленьких тельцах, обильно снабжаемых артериальной кровью. Важными для регуляции дыхания являются каротид-ные хеморецепторы. Аортальные хеморецепторы на дыхание влияют слабо и имеют большее значение для регуляции кровообращения. [28]
Эффект Пастера, изученный на яблоках Блекменом и Парийя [4], по мнению многих физиологов растений, проявляется при ресинтезе углеводов из органических кислот. Вычисления отношения [ АТФШАДФ1, необходимого для заметного снижения скорости гликолиза, дали значения, приблизительно в 1000 раз большие, чем наблюдавшиеся в действительности. Следовательно, отношение [ АТФ ] / [ АДФ ] не может осуществлять эффективную регуляцию дыхания. [29]
После отделения головного мозга от спинного на уровне верхних шейных сегментов дыхательные движения прекращаются. Лишь изредка удается наблюдать слабые сокращения дыхательных мышц, но они имеют неправильные ритм и форму. Если перерезать мозг на уровне нижних шейных сегментов, дыхательная активность диафрагмы сохраняется, а межреберных мышц - прекращается. Следовательно, в регуляции дыхания принимают участие и центры головного мозга. [30]
Страницы: 1 2 3