Регуляция - биосинтез - белок
Cтраница 1
Регуляция биосинтеза белка, протекающего с исключительно высокой ( до 100 пептидных связей в секунду. Информация, необходимая для биосинтеза ферментов, содержится в трех структурных генах, которые вместе с ответственным за транскрипцию операторным геном образуют единый комплекс - оперон. Индуктор действует через ранее включенный регуляторный ген на операторный ген. В отсутствие лактозы репрессор ( аллостерический белок) вступает во взаимодействие с регуляторным геном и таким образом блокированием всего опе-рона прекращает синтез ферментов. [1]
Мы рассмотрим теперь регуляцию биосинтеза белка как в качественно, так и в количественном аспекте. Иными с i вами, мы попытаемся ответить на вопрос, какие факторы определяют, сколько именно данного белка должно синтезироваться в тот или иной момент жизни клетки. При рассмотрении этого вопроса нам придется ограничиться в основном данными, касающимися бактериальных систем, поскольку факторы, регулирующие синтез белка, изучались преимущественно на этих системах. [2]
 |
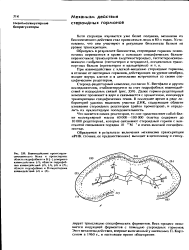
Схема оперона. [3] |
Описанный способ контроля и регуляции биосинтеза белка у прокариот еще не может обеспечить регуляторные нужды клетки. Белки, кодируемые одним и тем же опероном, могут требоваться в разных количествах и в разное время. Для понимания соответствующих регуляторных явлений необходимо детальное рассмотрение процесса транскрипции. [4]
Описанный способ контроля и регуляции биосинтеза белка у прокариотов еще не может обеспечить регуляторные нужды клетки. Белки, кодируемые одним и тем же опероном, могут требоваться в разных количествах и в разное время. Процесс образования начала молекулы РНК существенно отличен от последующего роста ее цепи. Соответственно рационально рассматривать начало синтеза РНК ( инициацию) и продолжение этого синтеза ( элонгацию) как две самостоятельные стадии. [5]
Естественно, что метаболическая стабильность ( долгоживучесть) эукариотической мРНК делает регуляцию на уровне трансляции особенно важной в общей картине регуляции биосинтеза белка. Например, показано, что инъекция женского полового гормона ( эстрогена) цыплятам индуцирует цитодифференцировку яйцеводов, синтез мРНК яичного альбумина и синтез этого белка. После прекращения инъекций синтез мРНК и синтез альбумина прекращаются, но можно показать, что предсинтези-рованная альбуминовая мРНК частично сохраняется в цитоплазме в нетранслируемой или слабо транслируемой форме. Повторная инъекция гормона ( эстрогена или прогестерона) немедленно реактивирует ее трансляцию, стимулируя инициацию. [6]
В тех случаях, когда нарушение структуры ДНК ( или РНК) под действием мутагенов произошло в участках, которые принимают участие в процессах регуляции биосинтеза белков, отмечаются изменения белкового синтеза всего организма. [7]
Исследования по биосинтезу белка быстро и непрерывно расширяются, и в этом кратком очерке мы даже не пытались осветить этот вопрос полностью; однако сказанного вполне достаточно, чтобы читатель получил основу для знакомства с проблемой регуляции биосинтеза белка. [8]
Нуклеиново-белковое взаимодействие имеет важнейшее-значение для регуляции биосинтеза белка, для регуляции действия генов. [9]
 |
Взаимодействие прсгестерон. [10] |
Хотя стероиды изучаются уже более полувека, механизм их биологического действия стал проясняться лишь в 80 - х годах. Установлено, что они участвуют в регуляции биосинтеза белков на уровне транскрипции. [11]
Можно считать установленным, что в основе таких разнообразных явлений, как взаимодействие антиген - антитело, рецепция запаха, регуляция биосинтеза белка в прокариотах и эукариотах, а также во взаимодействиях клеток лежат в принципе сходные механизмы узнавания. Именно узнавание на внутримолекулярном, межмолекулярном и надмолекулярном уровнях должно рассматриваться как молекулярная основа биофизики. Особо важную роль здесь играет матричный синтез. [12]
 |
Сверхспирализация двухцепочечной кольцевой ДНК под действием ДНК - гиразы. 1 - кольцевая форма ДНК. 2-сзерхспирализованная форма.| Участок РНК бактериофага. [13] |
Рибосомные рибонуклеиновые кислоты, связываясь с рибосомными белками, образуют рибосомы, в к-рых осуществляется синтез белка. Обнаружены т.наз. малые ядерные РНК, участвующие в превращ. РНК участвуют в регуляции биосинтеза белка и репликации плазмидных ДНК. В виде РНК представлены геномы мн. РНК-содержащие вирусы), в к-рых матрицами для синтеза РНК служат вирусные РНК. Нек-рыс РНК обладают ферментативной активностью, катализируя расщепление и образование фосфодиэфирных связей в своих собственных или др. молекулах РНК. [14]
Явления молекулярного узнавания, реализуемые посредством слабых взаимодействий, ответственны за совокупность регуляторных процессов в живой клетке, определяющих весь ход ее развития ( см. гл. Молекулярные взаимодействия формируют необходимые для регуляторных процессов обратные связи. Это относится, в частности, к функционированию аллостериче-ских ферментов ( [1], гл. Здесь мы рассмотрим молекулярную регуляцию биосинтеза белка, регуляцию действия генов. [15]
Страницы: 1