Генетическая рекомбинация
Cтраница 2
Вероятно, процесс эволюции благоприятствовал половому размножению, так как случайная генетическая рекомбинация увеличивала шансы организмов на то, что хотя бы некоторые из их потомков выживут в непредсказуемо изменчивом окружающем мире. В то же время половой процесс может облегчать распространение полезных мутаций в большой популяции. Половой процесс необходим также для поддержания диплоидности и таким образом способствует созданию условий для быстрой выработки новых генов у высших растений и животных. [16]
В основе одного из первых механизмов, предложенных для объяснения генетической рекомбинации, лежало предположение, что рекомбинация непосредственно связана с синтезом ДНК. ДНК ДО какой-то случайной точки, в которой полимераза перескакивает на вторую из двух гомологичных хромосом и начинает копировать ее. [17]
Другой вывод, относящийся к рекомбинантам - точное соблюдение закона генетической рекомбинации. [18]
Гораздо большее значение, чем мутации, для изменчивости видов имеют генетические рекомбинации. Они связаны с ассоциацией родительских молекул ДНК и появлением гибридных, дочерних макромолекул. [19]
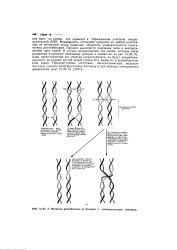 |
А. Механизм рекомбинации по Холидею с одиоцепочечиыми обменами. [20] |
Возможность остановки процесса на любом расстоянии от начальной точки позволяет объяснить универсальность генетических рекомбинаций. [21]
Садовод постоянно ищет отдельные садовые растения ( особи), которые благодаря резкой генетической рекомбинации или мутационным изменениям могут оказаться очень ценными. [22]
Рассмотрим теперь вкратце не совсем понятные химические явления, лежащие в основе таких явлений, как генетическая рекомбинация, интеграция вирусной ДНК с геномом клетки-хозяина и исключение про-фага из хромосомы клетки-хозяина. Бактерии с мутациями в некоторых из этих генов необычайно чувствительны к ультрафиолетовому облучению, что свидетельствует об их неспособности репарировать ( восстанавливать) повреждения ДНК, вызванные действием ультрафиолета ( гл. Из этого следует, что некоторые из ферментов, обеспечивающих процесс рекомбинации, нужны клетке также и для восстановления повреждений, вызванных действием ультрафиолетового излучения. Однако специфические функции большинства продуктов этих генов все еще до конца не выяснены. В геноме фага X имеются гены, кодирующие другую рекомбинационную систему, функционирующую независимо от продуктов генов фага A, inf и xis ( рис. 15 - 15), необходимых для интеграции и исключения генетического материала вируса и обеспечивающих процессы сайт-специфической ( для определенных участков геномов) рекомбинации между генами клетки-хозяина и вируса. [23]
Необходимо было доказать, что в культурах клеток F присутствуют мутанты Hfr и что именно они и ответственны за генетическую рекомбинацию. Напомним, что причина различия в том, что в разных культурах мутанты появляются в разное время и потому успевают размножиться в различной степени. [24]
Теперь, когда процесс конъюгации доказан для многих родов и видов бактерий, можно надеяться на успех использования его в целях изучения генетических рекомбинаций и для получения практически ценных штаммов - продуцентов аминокислот, витаминов и других веществ, а также для производства вакцин. [25]
Нормальный биологический обмен между генами или объединение генов из разных источников с образованием измененной хромосомы, способной после этого реплицироваться, транскрибироваться и транслироваться, называется генетической рекомбинацией. Она встречается в различных биологических ситуациях. Напомним, что в этом эксперименте ДНК из вирулентного штамма пневмококка попадала в клетки невирулентного штамма и превращала этот штамм в вирулентный. Очевидно, ген вирулентности, присутствующий в ДНК донорной клетки, включается в геном ре-ципиентной клетки. Такая трансформация бактериальных клеток, реализуемая вследствие рекомбинации генов, может наблюдаться не только в лаборатории, но и в естественных условиях. [26]
Таким образом, у диплоидного вида с половым размножением могут возникать новые гены в результате мутаций в добавочных копиях имеющихся генов; эти новые гены могут распространиться в популяции благодаря отбору в пользу гетерозигот, причем не будут потеряны и исходные гены; и наконец, новые гены могут дополнительно включаться в геном в результате процессов дупликации генов и генетической рекомбинации. Такая последовательность событий возмножна только у диплоидных видов. Обогащение генома у гаплоидного вида связано с большими трудностями. Если в процессе приобретения нового гена вид должен сохранить и старый ген, то ему придется ждать возникновения нужной мутации у одной из очень немногих особей, у которых уже произошла дупликация соответствующего локуса. Детальные расчеты показывают, что в типичном случае диплоидный организм способен расширять свой геном и добавлять к нему новые гены с новыми функциями в сотни или даже тысячи раз быстрее, чем это происходит у гаплоидного организма. [27]
Теперь мы можем отыскать спонтанные или индуцированные акридинами ревертанты к дикому типу, исходя из супрессорных мутантов. Снова путем генетической рекомбинации с диким фагом убеждаемся в том, что возврат к дикому типу произошел благодаря образованию второй мутации ( супрессора к супрес-сору, на этот раз со знаком) недалеко от первой. Процесс можно продолжать как угодно долго, все время отыскивая супрессорные мутанты все более высокого порядка. В итоге получается путем отбора спонтанных ревертантов и под; действием акридинов около 80 точечных мутантов. Все они по фенотипу были г, но их можно было разбить однозначно на две группы, приписав одной знак, другой - , в соответствии с чи - слом этапов при их получении. [28]
При генетических рекомбинациях новых генов в генофонде, как правило, не появляется. [29]
Еще до того как была окончательно установлена триплетная природа кодонов, Крик и его сотрудники, остроумно использовав мутации со сдвигом рамки, доказали, что генетический код действительно составлен из нуклеотидных триплетов. В результате генетической рекомбинации могут образоваться мутанты, содержащие обе мутации со сдвигом рамки. Однако распознать такие рекомбинанты будет трудно, так как ( согласно практически любой теории кодирования) они по-прежнему будут продуцировать полностью дефектные белки. Крику и его сотрудникам удалось, однако, ввести в тот же ген третью мутацию со сдвигом рамки того же типа и наблюдать, что рекомбинанты, несущие все три делеции ( или вставки), были способны синтезировать, по крайней мере частично, активные белки. Делеции одного или двух нуклеотидов полностью инактивируют ген, тогда как при делеции трех нуклеотидов, расположенных в пределах одного гена и близко друг от друга, ген укорачивается лишь на три нуклеотида. [30]
Страницы: 1 2 3 4