Репликация - плазмида
Cтраница 1
Репликация плазмид происходит независимо от репликации хромосомы, причем на одну хромосому в бактерии может приходиться одна или несколько одинаковых плазмид. Эписомы - это плазмиды, способные включаться в хромосому бактерии. Некоторые внехромосомные элементы могут вести себя как эписомы в одних бактериях и как плазмиды в других. Плазмиды могут быть инфекционными ( переносимыми) или неинфекционными. В первом случае они содержат гены для синтеза - половых пилей ( разд. А, 1 г) и способны - переносить свою ДНК в другую клетку. Если плазмида способна интегрироваться с хромосомой, а потом и выходить из нее, захватывая с собой при этом другие гены, то такую плазмиду называют фактором пола. Выше уже шла речь о том, что процесс переноса генов с помощью фактора F был широко использован при картировании бактериальных хромосом. [1]
Для своей репликации плазмиды используют репликативную машину клетки-хозяина, однако репликация плазмид происходит независимо от хромосомы. Каждая плазмида является самостоятельным репликоном, сама контролирует собственную репликацию и поддерживается в клетке в опредапенном, характерном для нее числе копий. Для характеристики плазмидных репликонов их принято разбивать на группы несовместимости. [2]
Для своей репликации плазмиды используют решшкативную машину клетки-хозяина, однако репликация плазмид происходит независимо от хромосомы. Каждая плазмида является самостоятельным репликоном, сама контролирует собственную репликацию и поддерживается в клетке в определенном, характерном для нее числе копий. Для характеристики плазмидных репликонов их принято разбивать на группы несовместимости. [3]
Для своей репликации плазмиды используют репликативную машину клетки-хозяина, однако репликация плазмид происходит независимо от хромосомы. Каждая плазмида является самостоятельным репликоном, сама контролирует собственную репликацию и поддерживается в клетке в определенном, характерном для нее числе копий. Для характеристики плазмидных репликонов их принято разбивать на группы несовместимости. [4]
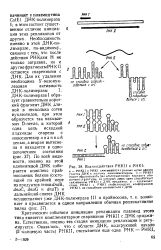
Критическим событием инициации репликации плазмид этого типа является комплементарное спаривание РНКИ с ДНК ориджи-на. Естественно, именно эта стадия инициации репликации и регулируется. [6]
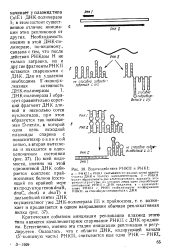 |
Взаимодействие РНКП и PHKI. [7] |
Критическим событием инициации репликации плазмид этого типа является комплементарное спаривание РНКП с ДНКориджи-на. Естественно, именно эта стадия инициации репликации и регулируется. [8]
Для своей репликации плазмиды используют репликативную машину клетки-хозяина, однако репликация плазмид происходит независимо от хромосомы. Каждая плазмида является самостоятельным репликоном, сама контролирует собственную репликацию и поддерживается в клетке в опредапенном, характерном для нее числе копий. Для характеристики плазмидных репликонов их принято разбивать на группы несовместимости. [9]
Для своей репликации плазмиды используют решшкативную машину клетки-хозяина, однако репликация плазмид происходит независимо от хромосомы. Каждая плазмида является самостоятельным репликоном, сама контролирует собственную репликацию и поддерживается в клетке в определенном, характерном для нее числе копий. Для характеристики плазмидных репликонов их принято разбивать на группы несовместимости. [10]
Для своей репликации плазмиды используют репликативную машину клетки-хозяина, однако репликация плазмид происходит независимо от хромосомы. Каждая плазмида является самостоятельным репликоном, сама контролирует собственную репликацию и поддерживается в клетке в определенном, характерном для нее числе копий. Для характеристики плазмидных репликонов их принято разбивать на группы несовместимости. [11]

Размеры плазмид варьируют от менее 1 до более 500 т.п.н. Каждая из них содержит сайт начала репликации ( ori), без которого репликация плазмиды в клетке-хозяине была бы невозможна. [13]
Представляет собой кольцевую, двунитевую молекулу ДНК, гены которой кодируют дополнительные свойства, придавая селективные преимущества клеткам. За счет автономной репликации плазмиды могут давать явление амплификации: одна и та же плазми-да. [14]
Изучению механизмов контроля репликации и числа копий: бактериальных плазмид посвящено множество работ. Было-обнаружено, что число копий плазмид примерно одинаково в клетках, размножающихся с разной скоростью. Из этого следует, что репликация плазмид как-то связана с ростом бактерий. Такая координация достигается при участии механизмов, контролирующих начало репликации. Если уж репликация началась, то она идет с относительно постоянной скоростью при любом темпе размножения бактерий. При высокой скорости размножения репликация индуцируется чаще в случае много-копийных плазм ид, чем малокопийных. [15]
Страницы: 1 2