Рецептор
Cтраница 3
Рецептор и ые геометрические модели в своей основе имеют приближенное представление геометрического объекта в плоскости или пространстве рецепторов. В области рецепторов строится прямоугольная решетка или сеть. [31]
Рецепторы нейромедиаторов исключительно важны для молекулярной нейробиологии, так как они играют ключевую роль при переносе нервных импульсов и являются центрами важных регуляторных процессов и тех изменений, которые происходят при некоторых нервных заболеваниях. Поскольку такие рецепторы представляют собой мишень действия многих нейрофар-макологических препаратов, они представляют как практический, так и теоретический интерес. Поэтому биохимия рецептор-ных молекул - одно из наиболее активно развивающихся сейчас направлений нейрохимических исследований. [32]
Адренэргические рецепторы в принципе не отличаются от а-рецепторов, но эффекторы действуют на них с другой активностью. Адреналин и норадреналин являются аго-нистами - рецепторов и почти так же эффективны для них, как для а-рецепторов. [33]
Рецепторы серотонина ( 5-гидрокситриптаминовые; или 5 - НТ-рецепторы) также действуют как множественные. [34]
Ацетилхоли-новые рецепторы в не иннервированных миотрубочках также распределяются неравномерно с образованием кластеров, которые детектируются оптическими методами с помощью а-бунга-ротоксина, меченного флуоресцентным красителем, или ауто-радиографически с использованием радиоактивно меченного токсина. Эти рецепторы практически иммобилизованы в кластерах, в то время как свободные рецепторы в мембране способны диффундировать и передвигаться. Естественно предположить, что кластеры образуют на поверхности клеток мышц области-мишени для врастающих нейритов, но, видимо, на самом деле происходит обратное: на участке, где нервное волокно впервые контактирует с мышечной клеткой, существенно повышается концентрация рецептора. Это обнаружено на смешанной культуре нейронов спинного мозга и клеток мышц лягушки Xenopus. Недавно было установлено, что в этом процессе принимает участие фактор белковой природы, способствующий агрегации. [35]
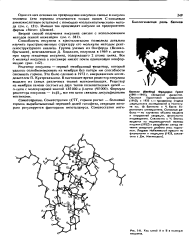 |
Ход цепей А к В в молеку к инсулина. [36] |
Рецептор инсулина - первый мембранный акцептор, который удалось солюбилизировать из мембран без потери им способности связывать гормон. [37]
Природные рецепторы главным образом должны - содержать оптически активные белковые компоненты, которые, как можно ожидать, отличают компетентный триггер от его энантиоморфа. [38]
Инсулино-вый рецептор подобен рецепторам многих ростовых веществ, т.е. обладает тирозинкиназной активностью. Отметим, что более 10 продуктов онкогенов, участвующих в пролиферации клеток, являются тирозинкиназами. [39]
Рецепторы вкуса - вкусовые почки - расположены на языке, задней стенке глотки, мягком небе, миндалинах и надгортаннике. [40]
Рецептор инсулина состоит из четырех полипептидных цепей двух типов и имее ф-лу а2р2; полипептидные цепи сшиты диеульфидными связями. В молекуле рецептора инсулина встречаются последовательности, гомологичные последовательностям рецепторов фа гороя роста. [41]
Рецепторы беспозвоночных устроены иначе, они во многих случаях чувствительны в широкой спектральной области. [42]
Шаровидные рецепторы фага Т5, которые были выделены из клеточной стенки бактерий. Кончик каждой фаго вой частиЧм блокирован шариком рецептора. [43]
Рецепторы стероидных гормонов представляют собой полипептиды с мол. В их структуре можно выделить домен, богатый остатками цистеина и основных аминокислот и ответственный за связывание с ДНК, и гидрофобный домен, ответственный за связывание стероидного гормона. Рецепторы половых стероидных гормонов гомологичны рецепторам глюкокортикоидов. [44]
Рецепторы второго типа находятся в суставах и посылают в мозг непрерывные сигналы о взаимном расположении различных частей тела. [45]
Страницы: 1 2 3 4