Ризогенез
Cтраница 1
Ризогенез стеблевых черенков представляет собой удобную модель для изучения индукционных процессов - образования новых тканей и целых органов. Клетки тканей стебля под действием эндогенных фитогормонов как бы перестраивают направление своего метаболизма, и в них начинается биосинтез новых продуктов, ведущих к заложению корневых зачатков. Дифференциация новых органов не ограничивается действием одних фитогормонов. [1]
Итак, цепь ризогенеза: индуктор-рецептор - первичные реакции-дедифференциация-дифференциация-органогенез имеет очень много неясных мест и нуждается в дальнейшем изучении. Этот процесс удобно исследовать на укореняющихся стеблевых черенках. [2]
Микроскопический контроль за этапами ризогенеза показал, что клеточные деления у основания стебля черенка обнаруживаются в зоне перицикла через два дня после начала укоренения. Затем этот процесс интенсивного деления распространяется и в камбиальную зону. ИУК резко активирует процесс заложения корневых зачатков. [3]
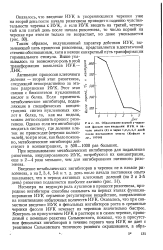 |
Образование корней у черенков фасоли при введении ИУК в начале опыта ( 0 и через 1 2 3 4 5 дней после постановки опыта ( Кефели и др., 19706. [4] |
При использовании метаболических ингибиторов для подавления ризогенеза, индуцированного ИУК, потребуются их концентрации, еще в 2 - 4 раза меньшие, чем для ингибирования нативного ризогенеза. [5]
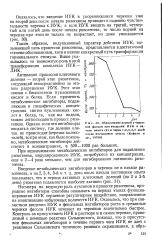 |
Образование корней у черенков фасоли при введении ИУК в начале опыта ( 0 и через 1 2 3 4 5 дней после постановки опыта ( Кефели и др., 19706. [6] |
Активация процессов клеточного деления - второй этап ризогенеза, следующий непосредственно за этапом разрушения ИУК. Этот этап связан с биосинтезом нуклеиновых кислот и белка. Если применить метаболические ингибиторы, подавляющие в специфических концентрациях синтез нуклеиновых кислот и белка, то, как мы видели, можно легко подавить процесс ризогенеза. Однако если эти ингибиторы вводить в систему, где деления клеток не происходит ( отрезки колеоп-тилей), то при этом, как было показано, метаболические ингибиторы действуют в концентрациях, в 500 - 1000 раз больших. [7]
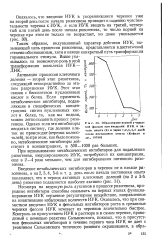 |
Образование корней у черенков фасоли при введении ИУК в начале опыта ( 0 и через 1 2 3 4 5 дней после постановки опыта ( Кефели и др., 19706. [8] |
Несмотря на ведущую роль ауксинов в процессе ризогенеза, природные ингибиторы способны оказывать на первых этапах функционирования ИУК серьезное регуляторное воздействие, направленное на инактивацию избыточных количеств ИУК. Оказалось, что при совместном введении ИУК и фенольных ингибиторов роста в срезанные черенки фасоли ИУК исчезает из черенков значительно быстрее. [9]
На основании приведенных результатов складывается общее впечатление об этапах ризогенеза и о системах, в нем участвующих. [10]
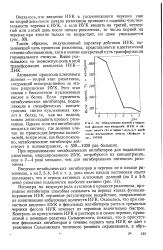 |
Образование корней у черенков фасоли при введении ИУК в начале опыта ( 0 и через 1 2 3 4 5 дней после постановки опыта ( Кефели и др., 19706. [11] |
Таким образом, индукционный характер действия ИУК, вызывающей цепь процессов ризогенеза, представляется в достаточной степени обоснованным, хотя и неясен конкретный путь трансформации гормонального стимула. [12]
Вместе с тем не остается постоянной и активность природных ингибиторов, которые осложняют процесс ризогенеза, хотя здесь не наблюдается такой четкой закономерности, как для ауксинов. Было показано также, что у черенков трудно укореняемых пород растений практически не происходит изменения ауксинов и ингибиторов в процессе укоренения, в то время как у легко укореняемых активность этих групп веществ резко изменяется. [13]
В отличие от фенольных ингибиторов абсцизовая кислота подавляет растяжение клеток отрезков колеоптилей в низких концентрациях, а процесс ризогенеза не ингибирует совсем. Абсцизовая кислота способна ингибировать рост семядолей и поступление воды в прорастающие семена яблони, а также задерживать синтез хлорофилла в таких семядолях. [14]
Хотя проблема регуляции корнеобразования ауксинами активно разрабатывается, она не исчерпывает всех вопросов, связанных с эндогенными факторами ризогенеза. Начиная с исследований Буйе-на и Вента ( Bouillenne, Went, 1933), фактор, индуцирующий образование корней и отличающийся по своим свойствам от ауксинов, получил название ризокалина. Согласно более поздним данным ( Bouillenne, Warland, 1947, 1953), гипотеза о присутствии в растении ризокалина состоит в следующем: 1) мобильный фактор синтезируется в листе и транспортируется в зону корнеобразования вместе с ауксинами; 2) в некоторых клетках этой зоны локализуется второй катализирующий фактор, который, объединяясь вместе с первым и с ауксином, индуцирует процесс корнеобразования. [15]
Страницы: 1 2 3