Рост - слоевище
Cтраница 1
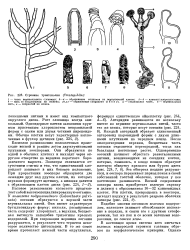 |
Строение трентеполии ( Trentepohlia. [1] |
Рост слоевища всегда апикальный. Одноядерные клетки заполнены крупным пристенным хлоропластом неправильной формы с одним или двумя четкими пиреноидами. [2]
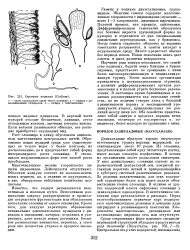 |
Строение кодиума ( Codium. [3] |
Рост слоевища в длину обусловлен апикальным вытягиванием центральных нитей. Образование новых пузырей среди уже существующих не только ведет к более плотному их расположению, но и представляет собой форму интеркалярного роста слоевища. У стелющихся подушковидных форм этот способ роста преобладает. [4]
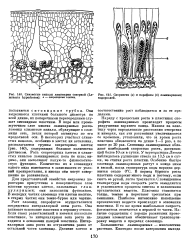 |
Слизистые каналы ламинарии северной ( La-minaria hyperborea. - секреторные клетки.| Спорангии ( а и парафизы ( б ламинариевых водорослей. [5] |
Рост слоевищ спорофитов осуществляется посредством интеркалярной зоны роста. Она занимает основание пластины и вершину ствола. Если ствол разветвленный и имеется несколько пластинок, то интеркалярная зона роста находится в основании каждой пластины. Интеркалярная зона роста не отграничена резко от остальной части слоевища. [6]
Скорость роста слоевища лишайников существенно зависит от общих экологических условий - температуры, осадков, влажности воздуха. На крупных территориях со сравнительно одинаковым климатом она может оставаться более или менее одинаковой. [7]
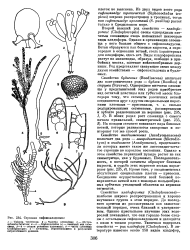 |
Строение сифонокладовых. [8] |
В обоих родах рост слоевища с самого начала правильный, симметричный ( рис. 235, 3), На каждом сегменте возникает пара боковых ветвей, которые развиваются синхронно и повторяют тот же способ роста. [9]
 |
Риелла ( Riella - мужское и женское растения.| Моноклея ( Monoclea. [10] |
Архегонии закладываются позади верхушечной клетки, и после их образования рост слоевища не прекращается. Архегонии часто расположены в специальных образованиях на спинной стороне слоевища или на специальных ветвях, а у листостебельных видов - вдоль средней линии спинной стороны стебля. Аналогично расположение и антериди-ев. Основание спорогона защищено псевдопери-антием, или колпачком, или тем и другим. Степки коробочки 2 - или 3 - 6-слойные, с утолщениями клеточных стенок или без них. Порядок включает около 10 семейств. [11]
В отличие от накипных и листоватых форм лишайников, для которых характерен горизонтальный рост гиф, у кустистых лишайников наблюдается вертикально направленный рост гиф и верхушечный рост слоевищ. Это позволяет кустистым лишайникам путем изгибов веточек в разные стороны занимать наилучшее положение, при котором водоросли могут максимально использовать свет для осуществления фотосинтеза. Эти лишайники обычно прикрепляются к субстрату только небольшим участком нижней части слоевища. Прямостоячие напочвенные кустистые лишайники чаще всего прикрепляются к почве тонкими нитевидными ризоидами. Прикрепление повисающих слоевищ кустистых лишайников к коре дерева или поверхности скал происходит с помощью п сев-до г о м ф а. Последний имеет вид короткой ножки, расширенной на конце в небольшую пяточку ( рис. 293); он напоминает по внешнему виду гомф листоватых лишайников, но отличается от него анатомическим строением. Слоевища кустистых лишайников могут быть разных размеров. [12]
Иными словами, бентосные водоросли, растущие в условиях движения воды, получают преимущества по сравнению с водорослями в тихих местах: один и тот же уровень фотосинтеза может быть достигнут у них при меньшей освещенности, что в итоге способствует росту более крупных слоевищ с большим содержанием белка и углеводов. Движение воды предотвращает оседание на скалы и камни илистых частиц, которые мешают закреплению зачатков водорослей. Кроме того, движение воды благоприятствует росту бентосных водорослей тем, что с поверхности грунта удаляются животные, питающиеся водорослями. Наконец, хотя при сильном течении или сильном прибое происходит повреждение слоевищ водорослей или отрыв их от грунта, движение воды все же не препятствует поселению микроскопических водорослей и микроскопических стадий крупных водорослей. Дело в том, что из-за трения о поверхность грунта скорость движения воды по мере приближения к нему уменьшается и на самой поверхности грунта существует так называемый пограничный слой воды, где течение отсутствует. Толщина этого слоя прямо пропорциональна протяженности обтекаемого предмета ( например, камня или выступа скалы) и обратно пропорциональна скорости течения. Обычно толщина пограничного слоя бывает не меньше 10 - 100 мкм, что соответствует размерам спор, гамет и зигот большинства бентосных водорослей. [13]
В том случае, когда на поверхности слоевища лишайников не образуются специальные органы газообмена, на помощь приходят различные трещинки и разрывы в коровом слое. Эти трещинки могут возникнуть как в результате особенностей роста слоевища, так и в результате повреждения слоевища ветром, животными, человеком. [14]
Представителям этого порядка свойственна регулярная гетероморфная смена форм развития. В отличие от кутлериевых волоски у спо-рохновых не принимают участия в росте слоевища. У сильно разветвленных представителей пучки волосков играют существенную роль в фотосинтезировании органических веществ. Поэтому у видов родов карпомитра и спорохнус спорангии располагаются вблизи вершин ветвей. [15]
Страницы: 1 2