Сверхспирализация
Cтраница 3
Поли ( / - - глутаминовая кислота) подобно другим полиаминокислотам, имеющим ионизуемые боковые группы, легко растворима в воде и при значениях рН, достаточно низких, чтобы подавить - ионизацию, принимает конформацию сс-спирали. Дополнительная спирализация, образующаяся за счет сетки связанных водородными связями у-карбоксильных групп, еще больше стабилизует структуру. Такой сверхспирализации способствует добавление к раствору диоксана или гидроксилсодержащих растворителей. [31]
Изучение сверхспирализации ДНК, замкнутых in vitro с помощью ДНК-лигазы 315 316 331 332, подтверждает это положение. Очень высокую степень сверхспирализации удается получить 316, если замыкание цикла проводить в присутствии интерка-лирующих красителей ( вызывающих частичное расплетание двойной спирали ДНК), а после циклизации удалить их из молекулы ДНК. Исследование сверхспирализации ковалентно-замкнутых ДНК, выделенных из различных источников, может пролить свет на условия их образования m vivo. В качестве меры степени сверхспирализации ДНК можно принять число витков сверхспирали, приходящееся на единицу длины молекулы. Иногда используют величину а, называемую плотностью сверхспирали, которая является частным от деления числа сверхспиральных витков т на число р, равное / го общего числа нуклеотидов в изучаемой двух-спиральной молекуле. [32]
Какие изменения в структуре ДНК может вызывать сверхспирализация. Понятно, что выгодным будет любое изменение структуры, в результате которого произойдет ослабление напряжения, вызванного в зкДНК сверхспира-лизацией. Поэтому было ясно, что сверхспирализация должна способствовать образованию в двойной спирали раскрытых областей, а также крестообразных структур. [33]
Открытие топоизомераз заставило усомниться в том, что сверхспирализация никчемна в биологическом смысле. Ведь, если есть ферменты, меняющие топологию, то, значит, сама топология клетке не совсем безразлична. [34]
Сверхспирализация - это важнейший пример того, как физическое состояние молекулы ДНК влияет на ее работу в клетке. Всю эту проблему интенсивно изучают специалисты самых разных профилей - от медиков до математиков. Поэтому неудивительно, что существует множество гипотез о роли сверхспирализации в работе клетки. Мы остановимся более подробно на одной из них, которая кажется сейчас наиболее простой и правдоподобной. [35]
Последнее утверждение может вызвать недоумение. Ведь кажется, что сверхспираль на рис. 23 правая, а не левая. Это один из парадоксов, с которыми приходится сталкиваться при изучении сверхспирализации. [36]
Развертывание двунитевой спиральной структуры геликазами не создает каких-либо осложнений, если, как это представлено на рис. 50, концы ДНК свободны. Если же они закреплены, как это, бесспорно, имеет место в кольцевых ДНК и скорее всего у ДНК в хромосомах эукариот, то раскручивание двойной спирали создает в остальной части структуры сверхспирализацию. При этом, поскольку раскручивается правая спираль, возникает и постепенно усиливается положительная сверхспирализация. Это не может не сказываться на протекании процессов в вилке репликации и должно постепенно привести к торможению процесса в целом. Чтобы избежать этого, необходимо введение в сохранившуюся двуспираль-ную структуру отрицательных супервитков. [37]
Хромосомная ДНК, как правило, сверхспирализована. Как это было впервые показано в лаборатории Георгиева в 1982 г. ( Лучник и Бакаев), сверхспирализация ДНК играет важную роль в биологической активности генома. Различные нуклеотид-ные последовательности в молекуле ДНК конкурируют за упругие витки и энергию сверхспирализации, поглощая их в конфор-мационных переходах. Конформационные изменения, связанные с этими напряжениями, имеют прямое значение для регуляции генов. Сверхспирализация генома изменяется при дифференцировке, старении и злокачественной трансформации клеток. [38]
 |
Такой крест об-разуется в ДНК ColEl, когда она находится в. [39] |
С ростом сверх-спирализации картина очень силь-но меняется. Быстро увеличивается вероятность образования раскры-тых пар. Но вероятность образо-вания крестов в коротких пере-вертышах вроде тех, которые уз-наются рестриктазами, остается пренебрежимо малой при любых значениях сверхспирализации. [40]
Гипотеза эта возникла потому, что было прямо доказано: для того чтобы начать удваиваться, молекуле ДНК обязательно надо закрутиться в сверхспираль, но для самого процесса репликации сверхспираль вовсе не нужна. Более того, иногда перед репликацией одна из нитей кольцевой замкнутой ДНК рвется, причем этот разрыв делает специальный белок и только в том случае, если ДНК сверх-спирализована. Получается какая-то бессмыслица - клетка затрачивает усилия, чтобы превратить ДНК в сверхспираль с помощью одного белка ( ДНК-гиразы) для того, чтоиы другой белок эту сверхспирализацию немедленно ликвидировал. [41]
Стало ясно, что сверхспнрализованное состояние ДНК не исключение, как думали вначале, а правило. Но тут возникло сомнение, - а такова ли ДНК там, внутри клетки. По-виднмому, сверхспирализация - это реакция на насильственное извлечение ДНК из родной стихии, ведь условия, в которых пребывает ДИК внутри клетки, конечно же, отличаются от условий после ее извлечения. [42]
Изучение сверхспирализации ДНК, замкнутых in vitro с помощью ДНК-лигазы 315 316 331 332, подтверждает это положение. Очень высокую степень сверхспирализации удается получить 316, если замыкание цикла проводить в присутствии интерка-лирующих красителей ( вызывающих частичное расплетание двойной спирали ДНК), а после циклизации удалить их из молекулы ДНК. Исследование сверхспирализации ковалентно-замкнутых ДНК, выделенных из различных источников, может пролить свет на условия их образования m vivo. В качестве меры степени сверхспирализации ДНК можно принять число витков сверхспирали, приходящееся на единицу длины молекулы. Иногда используют величину а, называемую плотностью сверхспирали, которая является частным от деления числа сверхспиральных витков т на число р, равное / го общего числа нуклеотидов в изучаемой двух-спиральной молекуле. [43]
Функцию раскручивания ( расплетения) двойной спирали ДНК в репли-кационной вилке, происходящего за счет энергии гидролиза АТФ, выполняет специфический rep - белок, названный хеликазой ( мол. Образовавшиеся на определенное время одноцепочечные участки ДНК служат в качестве матрицы при репликации и стабилизируются при помощи особых белков, связывающихся с одноцепочечной ДНК ( ДНК-связывающие белки) и препятствующих обратному комплементарному взаимодействию цепей ДНК ( мол. В связи с этим их иногда называют дестабилизирующими двойную спираль белками. Имеются, кроме того, особые ферменты топоизомеразы ( у прокариот одна из них названа ДНК-гиразой), которые играют особую роль в сверхспирализации, обеспечивая как репликацию, так и транскрипцию ДНК. Эти ферменты наделены способностью не только создавать супервитки, но и уничтожать суперспирализацию путем сшивания образующихся разрывов или разрезания ДНК. [44]
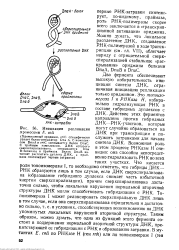 |
Инициация репликации хромосомы Е. coli. [45] |
Страницы: 1 2 3 4