Отрицательная сверхспирализация
Cтраница 1
Отрицательная сверхспирализация, наряду с крестообразными структурами, стабилизует и Z-форму ДНК. [1]
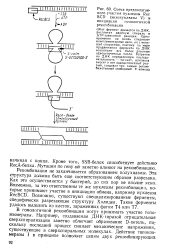 |
Схема предполагаемого участия нуклеазы Rec BCD ( экзонуклеазы V & инициации гомологичной рекомбинации. [2] |
В гомологичной рекомбинации могут принимать участие топо-изомеразы. Например, создаваемая ДНК-гиразой отрицательная сверхспирализация заметно облегчает образование D-петель, поскольку последний процесс снимает механические напряжения, существующие в сверхспиральных молекулах. [3]
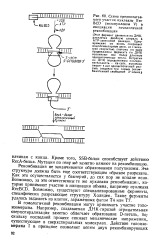 |
Схема предполагаемого участия нуклеазы Rec BCD ( экзонуклеазы V в инициации гомологичной рекомбинации. [4] |
В гомологичной рекомбинации могут принимать участие топо-изомеразы. Например, создаваемая ДНК-гиразой отрицательная сверхспирализация заметно облегчает образование D-петель, по-скольку последний процесс снимает механические напряжения, существующие в сверхспиральных молекулах. Действие топоизо-меразы I в принципе позволяет цепям двух рекомбинирующих. [5]
 |
Схема предполагаемого участия нуклеазы Rec BCD ( экзонуклеазы V з инициации гомологичной рекомбинации. [6] |
В гомологичной рекомбинации могут принимать участие топо-изомеразы. Например, создаваемая ДНК-гиразой отрицательная сверхспирализация заметно облегчает образование D-петель, по-скольку последний процесс снимает механические напряжения, существующие в сверхспиральных молекулах. [7]
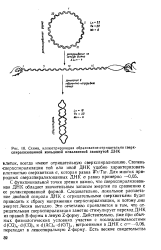 |
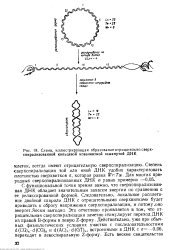
Схема, иллюстрирующая образование отрицательно сверх-спиралнэованной кольцевой ковалентной замкнутой ДНК. [8] |
С функциональной точки зрения важно, что сверхспирализован-ная ДНК обладает значительным запасом энергии по сравнению с ее релаксированной формой. Это отчетливо проявляется в том, что отрицательная сверхспирализация заметно стимулирует переход ДНК из правой В-формы в левую Z-форму. Действительно, уже при обычных физиологических условиях участки с последовательностями d ( CG) n - d ( CG) n и d ( AC) n - d ( GT) n, встроенными в ДНК с а-0 06, переходят в левоспиральную Z-форму. [9]
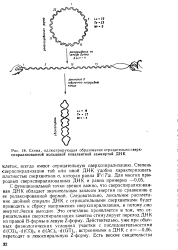 |
Схема, иллюстрирующая образование отрицательно сверх-спирализованной кольцевой кйвалентной замкнутой ДНК. [10] |
С функциональной точки зрения важно, что сверхспирализован-ная ДНК обладает значительным запасом энергии по сравнению с ее релаксированной формой. Это отчетливо проявляется в том, что отрицательная сверхспирализация заметно стимулирует переход ДНК из правой В-формы в левую Z-форму. Действительно, уже при обычных физиологических условиях участки с последовательностями d ( CG) n - d ( CG) n и d ( AC) n - d ( GT) u, встроенными в ДНК с а-0 06, переходят в левоспиральную Z-форму. [11]
 |
Схема, иллюстрирующая образование отрицательно сверх-спирализованной кольцевой ковалентной замкнутой ДНК. [12] |
С функциональной точки зрения важно, что сверхспирализован-ная ДНК обладает значительным запасом энергии по сравнению с ее релаксированной формой. Это отчетливо проявляется в том, что отрицательная сверхспирализация заметно стимулирует переход ДНК из правой В-формы в левую Z-форму. Действительно, уже при обычных физиологических условиях участки с последовательностями dCCG - dCCG) и d ( AC) n - d ( GT) l, встроенными в ДНК с 0 - 0 06, переходят в левоспиральную Z-форму. [13]
Все эти проблемы разрешаются прц. Способность бактериальной топоизомеразы II - ДНК-гиразы - отрицательно сверхспирализовать ДНК в АТР-зависимой реакции не только снимает вопрос о вращении всей непрореплицировавшейся части ДНК, но и облегчает действие хеликаз, поскольку отрицательная сверхспирализация, которую гираза создает перед вилкой, способствует расплетанию ДНК. [14]
Все эти проблемы разрешаются присутствием в клетке топоизомераз ( см. гл. Способность бактериальной топоизомеразы II - ДНК-гиразы - отрицательно сверхспирализовать ДНК в АТР-зависимой реакции не только снимает вопрос о вращении всей непрореплицировавшейся части ДНК, но и облегчает действие хеликаз, поскольку отрицательная сверхспирализация, которую гираза создает перед вилкой, способствует расплетанию ДНК. [15]
Страницы: 1 2