Септа
Cтраница 3
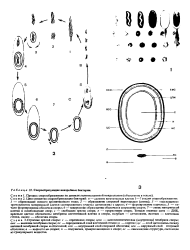 |
Спорообразующие анаэробные бактерии. [31] |
Цикл развития спорообразующих бактерий: в - деление вегетативных клеток 1 - 7 стадии спорообразования: / - образование осевого хроматинового тяжа; 2 - образование споровой перегородки ( септы); 3 - поглощение протопластом материнской клетки септированного участка цитоплазмы с ядром; 4 - формврование кортекса; 5 - начало формирования оболочки споры; 6 - завершение образования оболочки и созревание споры; 7 - лизис материнской клетки и освобождение спор; с - свободная зрелая спора; п - прорастание споры. Темные извитые нити - ДНК, красным цветом обозначены мембраны вегетативной клетки и споры, голубым - цитоплазма, желтым - клеточная стенка, синим - оболочка споры. [32]
ЯШСМ), дают начало восходящей системе ак сонов азующих а пути т к сток дорсального ядра преимущест-венн к ук ам полосатого тела и нейронов медиального ядра к структурам гипоталамуса, септа - иппокампальнои области, ядрам миндалевидного комплекса и коре пот шарий мозга. Имеются многочисленные данные об участии этой системы мозга в анксиолитических эффектах 4-бензо иазепинов. [33]
У некоторых диатомеи, имеющих вытянутые створки, на внутренней поверхности вставочных ободков образуются кремнеземные перегородки, или септы, вдающиеся в полость панциря параллельно плоскости створок. Септы возникают или по всей внутренней поверхности вставочного ободка, или только в одном из его концов. Они обычно хорошо видны со стороны пояска, различны по положению, форме и размерам и имеют одно или несколько отверстий. [34]
Гастральная полость разделена септами на 4 кармана. Каждая септа образована эктодермальной воронкой, сужающейся к основанию и содержащей продольный мускул-ретрактор. [35]
Некоторые органеллы клетки, даже ядра, могут мигрировать через поры. Наличие пор в септах играет существенную роль в переносе питательных веществ по гифам в зону роста. [36]
Итак, для образования перегородки и клеточного деления необходимы как завершение репликации ДНК, так и достижение пороговой длины клетки. Повреждение ДНК ингибирует образование септы. Для инициации образования перегородки важно наличие FtsZ-белка. Этот белок между делениями равномерно распределен в цитоплазме клетки, а в начале деления он формирует z - коль-цо в месте образования септы. Необходимо также участие белков, осуществляющих трансгликозилирование и транспептидацию в синтезе пептидогликана при создании новой клеточной стенки. [37]
Регенераторная способность гепатоцитов на начальных стадиях цирроза печени не только не ослабевает, но даже усиливается. Однако размножающиеся в замкнутых соединительнотканными септами пространствах клетки печени теперь уже не образуют тканевых структур, свойственных нормальной печеночной дольке, а располагаются беспорядочно, хаотически. [38]
На фермент-субстратные отношения влияет также изменение структуры ПГ, в частности его О, N - ацетилирование. Предполагают, что ПГ стенок септы после разделения клеток модифицируется и становится устойчивым к действию автолизинов. [39]
Вегетативное тело аскомицетов - разветвленный гаплоидный мицелий, состоящий из многоядерных или одноядерных клеток. В отличие от зигомицетов перегородки ( септы) в мицелии аскомицетов образуются упорядоченно, синхронно с делением ядер. Развитие септ происходит от стенок гифы к центру, напоминая сужение диафрагмы в объективе фотоаппарата. [40]
 |
Тонкое строение Neisseria catarrhalis. [41] |
Химический же состав его клеточных стенок отличается от такового у грамотрицательных бактерий. Механизм клеточного деления заключается в образовании септы, состоящей из ци-топлазматической мембраны и внутреннего слоя клеточной стенки. Внешняя мембрана при этом не втягивается вместе с внутренним слоем при образовании перегородки, а формируется заново при разделении клеток. Одновременно с образованием перегородки наблюдается и перешнуровывание. Исследователи, описавшие эту культуру, предложили такие грамвариа-бильные кокки, относимые ранее к микрококкам, поместить в отдельный род, промежуточный между истинными грамположительными и грамотрицательными бактериями. [42]
Цикл развития пчелиной аскосферы хорошо изучен. Мицелий гриба имеет типичные для аскомицетов септы, его клеточные стенки содержат хитин. Все виды аскосферовых гетеротал-личны, и у них наблюдается половой диморфизм ( морфологический гетероталлизм), необычный для аскомицетов. Мицелии разного пола хорошо различаются по окраске. При развитии совместимых мицелиев на одном из них образуются аскогоны с трихогинами, сливающимися с вегетативными клетками мицелия противоположного пола. Затем из аскогона развивается своеобразная спороносная структура, называемая спороцистом. Аскогон разрастается, протопласт внутри его одевается оболочкой и образует аскогенные гифы из двухъядерных клеток. На них но способу крючка образуются сумки, собранные в округлые группы. К моменту созревания аскоспор стенки отдельных сумок разрушаются, но их группы одеваются собственной тонкой оболочкой. Одновременно с развитием сумок оболочка аскогона хитинизируется и приобретает оливковую или темно-коричневую окраску. [43]
Двухнитевая ДНК спирализуется, скручивается в кольцо в точке прикрепления к цитоплазматической мембране. Это является сигналом для расхождения клеток по септе. Образуются две дочерние особи. [44]
Нарушение целостности клетки при автолизе начинается в местах преимущественной локализации автолитических ферментов, участвующих в процессах растворения участков клеточной стенки при росте и делении клетки. У грамположительных бактерий это происходит в месте образовании перегородки - септы, у гра-мотрицательных - в районе перетяжки. Здесь совершается синтез стеночных автолизинов, их активация, а также расположены участки стенки, пептидогликан которых имеет наибольшую субстратную чувствительность к автолизинам. [45]
Страницы: 1 2 3 4 5