Регуляторное взаимодействие
Cтраница 1
Детальный молекулярный механизм регуляторных взаимодействий при синтезе ферментов пока неясен, но описанная выше модель этого механизма создана на основе большого числа экспериментальных данных. Синтез ферментов происходит в рибосомах, и установлено, что состав рибосомальной РНК по своему нуклеотидному составу не является репликой ДНК. [1]
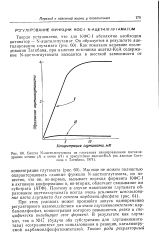 |
Синтез N-ацетилглутамата из глутамата изолированными митохондриями печени ( / и почек ( / / в присутствии ацетил - КоА ( по данным Сиге. [2] |
При этих условиях может произойти запуск одновременно регуляторного взаимодействия на этапе глутаминсинтетазы: кар-бамоилфосфат, концентрация которого повышается, резко и специфически ингибирует этот фермент. В результате как глута-мат, так и NH ( будучи оба субстратами для глутаминсинтетазы) сохраняются для использования в реакции, катализируемой карбамоилфосфатсинтетазой. [3]
У других организмов солевые АТФазы подвержены сильному модулирующему действию аденилатов, специфических фосфолипидов и, возможно, двухвалентных катионов, но в отношении рыб систематического изучения этих эффектов не проводилось. И наконец, активность ферментов слишком часто исследовали при неподходящей температуре ( 37 С); выбор столь небиологических условий может вести к совершенно неверным представлениям о фундаментальных регуляторных взаимодействиях. Очевидно, что все эти вопросы еще требуют изучения. [4]
 |
Схема транскрипции генома аденовируса. [5] |
Так, они регулируют эффективность считывания самого района Е1А: основной эффект заключается в стимуляции экспрессии этого района; другими словами, эти ранние гены как бы подстегивают сами себя. Однако при определенных условиях продукты района Е1А могут тормозить собственную работу. Природа этих сложных регуляторных взаимодействий еще не выяснена. [6]
В открытых системах могут устанавливаться простые обратные связи в силу сопряжения констант скоростей химических реакций внутри системы, а также констант переноса и обмена субстратов и продуктов с внешней средой. Эти обратные связи устанавливаются при участии значительных количеств вещества и энергии. Управление же на основе информации характеризуется избирательным контролем при помощи очень малых масс или энергий, так что энергетическая характеристика регуляторного взаимодействия отступает на второй план. [7]
Вторая важная особенность взаимодействий пируваткиназы Coryphaenoides с ФДФ состоит в том, что ФДФ снимает инги-бирующее действие АТФ только при высоком давлении. Необходимость высокого давления, без которого этот регуляторный эффект отсутствует, означает, что глубоководный фермент приспособлен для регулируемой функции только при высоких давлениях. В связи с этим возникает существенный вопрос относительно функции пируваткиназы у пелагических ювениль-ных форм, обитающих на меньшей глубине: выживают ли они с этим барофильным ферментом - возможно, благодаря каким-то другим регуляторным взаимодействиям, снимающим ин-гибирующий эффект АТФ. Или, может быть, на этой стадии развития они используют другой изофермент, приспособленный к меньшим давлениям. [8]
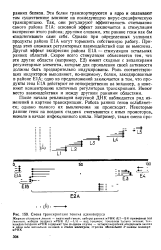 |
Схема транскрипции генома аденовируса. [9] |
Эти белки транспортируются в ядро и оказывают там существенное влияние на последующую вирус-специфическую транскрипцию. Так, они регулируют эффективность считывания самого района Е1А: основной эффект заключается в стимуляции экспрессии этого района; другими словами, эти ранние гены как бы подстегивают сами себя. Однако при определенных условиях продукты района Е1А могут тормозить собственную работу. Природа этих сложных регуляторных взаимодействий еще не выяснена. [10]
Одним из двух субстратов этого фермента является глутамин. Поэтому у брюхоногих моллюсков КФС и амидотранс-фераза непосредственно конкурируют за глутамин ( рис. 69), который у них и у большинства других животных играет роль переносчика аминогрупп. Так же как и КФС-I у позвоночных, КФС брюхоногих моллюсков активна только в присутствии N-ацетилглутамата. В условиях высокой концентрации глута-мата, характерной для тканей улиток, N-ацетилглутамат, вероятно, тоже должен образовываться в относительно больших количествах, а это должно вести к сильной специфической активации КФС и кратковременному накоплению карбамоилфосфата. Поскольку глутамин служит непосредственным субстратом для КФС, это регуляторное взаимодействие фактически приводило бы к переключению потока азота с пути синтеза мочевой кислоты на путь, ведущий к мочевине. [11]
Страницы: 1