Зооспора
Cтраница 2
 |
Циклы развития водорослей. [16] |
Фототаксис зооспор бентосных ( донных) водорослей, первоначально положительный, со временем сменяется на отрицательный, что обеспечивает оседание их на дно. Отрицательный фототаксис наблюдается также у зигот донных водорослей. Мужские гаметы обладают хемотаксисом, позволяющим им двигаться в сторону неоплодотворенных женских гамет, выделяющих особые химические вещества. Обнаружено, что споры некоторых бентосных морских водорослей, видимо, путем изменения объема тела и, таким образом, удельного веса концентрируются в слоях воды с определенной температурой и соленостью. В зависимости от направления течения в этих слоях споры выносятся к определенным участкам побережья, где и происходит развитие слоевищ. [17]
 |
Формы полового процесса у водорослей. [18] |
Споры и зооспоры обычно выходят в воду через отверстие в стенке спорангия целой группой, окруженные слизистой оболочкой, которая вскоре расплывается. Зооспоры по выходе из спорангия, находясь еще в общей оболочке, начинают активно двигаться и после разрыва оболочки моментально расплываются в разные стороны. Подобным образом выходят и гаметы при половом размножении. [19]
Перед образованием зооспор или гамет содержимое многогнездных вместилищ делится тонкими перегородками на камеры, в которых оказываются заключенными по одному ядру с участком цитоплазмы. В каждой камере развивается по одной, реже по две зооспоры или гаметы. [20]
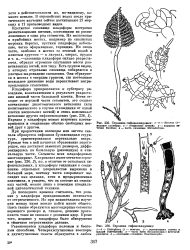
При прорастании зооспоры или зиготы сначала образуется сифонная булавовидная структура, ориентированная вертикально вверх. Прежде чем в ней начнется образование перегородок, она достигает заметных размеров, дифференцируясь на базальную ( ризоидную) и апикальную части. Сегменты всех кладофоровых многоядерные. Хлоропласт имеет сетчатое строение ( рис. 236, 2), но, в отличие от остальных сифонокладовых, у кладофоры тенденция к соединению отдельных хлоропластов выражена в большей мере, поэтому часто хлоропласт выглядит как цельная, хотя и продырявленная пластинка, и увидеть, что он состоит из отдельных частей, можно лишь с помощью специальной цитологической техники. [22]
 |
Ризоподии у некоторых Chrysophyta ( по Матвиенко, 1965. [23] |
После образования зооспор пробочка, закрывающая пору в-цисте, растворяется, и они выходят наружу. Форма и размеры цист постоянны для вида и служат систематическим признаком. Образуются цисты обычно при наступлении неблагоприятных внешних условий. [24]
Размножение осуществляется двужгутиковыми зооспорами, автоспорами, акинетами. Половой процесс изогамный или, у некоторых, оогамный. У одноклеточных представителей зооспоры или автоспоры, освободившись от оболочки материнской клетки, рассеиваются и вырастают каждая в новую особь, у колониальных они слагаются в новую колонию. [25]
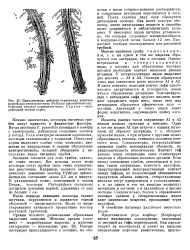 |
Привлекающее действие пораненных участков корней для зооспор пптиума ( Pythium aphanidermtitum. Стрелкой указано пораненное место. Справа - непораженный копчик корня. [26] |
Помимо химических, зооспоры питиевых грибов могут привлечь и физические факторы. Если электроды заменяли корешками, зооспоры скапливались у корешков. Поскольку корни выделяют слабые токи, возможно, движение по направлению к корням обусловлено электротаксисом, который обнаружен у нескольких видов питиевых грибов. [27]
 |
Привлекающее действие пораненных участков корней для зооспор пптиума ( Pythium aphanidermtitum. Стрелкой указано пораненное место. Справа - непораженный копчик корня. [28] |
После периода движения зооспоры теряют жгутики, округляются и одеваются тонкой оболочкой - инцистируются. Вслед за инци-стированием начинается прорастание зооспор ростковой трубочкой. [29]
Через некоторое время зооспоры оседают на то же слоевище или на другие водоросли и преобразуются в одноклеточные женские и мужские половые растения. В дальнейшем женские растения превращаются в оогоний с одной яйцеклеткой. В мужских растениях содержимое делится с образованием дву -, четы-рехжгутиковых антерозоидов. [30]
Страницы: 1 2 3 4 5