Скорость - трансформация
Cтраница 2
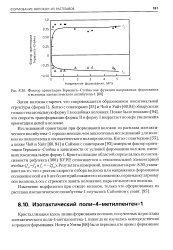 |
Фактор ориентации Германса-Стейна как функция напряжения формования в волокнах изотактического полибутена-1. [16] |
Позже было показано [94], что скорость трансформации формы II в форму I возрастает по мере увеличения ориентации цепей в волокне. [17]
Как правило, трансформации происходят между разными штаммами одного и того же вида. Было показано, что в таких случаях скорость трансформации зависит от количества частиц ДНК в трансформирующей среде. Известны также примеры трансформаций между разными видами бактерий, но скорость трансформации в этих случаях в несколько тысяч раз меньше, чем при трансформации внутри одного вида. [18]
К настоящему времени получены константы скорости реакции 1-го порядка практически для всех классов органических соединений; значения для некоторых из них приведены в табл. VI-4, а для наиболее изученных - фенолов - в табл. VI-5 и VI-6. Существенно, что эта простая модель ( по которой скорость трансформации вещества линейно зависит от его концентрации) формально согласуется с экспериментальными данными, даже когда скорость трансформации органического вещества в действительности не зависит от его концентрации, а определяется скоростью развития микроорганизмов. Однако это формальное соответствие характерно только для экспоненциальной фазы роста микроорганизмов; в период лаг-фазы и стационарной фазы их развития модель даже чисто формально не соответствует экспериментально наблюдаемой картине. Поэтому при обработке экспериментальных данных соответствующие этим фазам периоды трансформации веществ обычно не учитываются, и количественно оцениваются лишь не имеющие перегиба и плато участк кривых с экспоненциальным снижением концентрации веществ. Однако даже на этих участках кривых модель не адекватна, о чем свидетельствует изменение константы скорости во времени и ее зависимость от начальной концентрации вещества. Следовательно, эта модель и особенно полученные в экспериментах значения константы скорости не могут непосредственно использоваться для расчета скорости самоочищения вод в природных условиях. [19]
Приведенная методика анализа товарных запасов имеет особую значимость для торговых предприятий, в которых, как известно, основная часть оборотных активов приходится именно на товарные запасы. Выше уже упоминалось, что эффективность текущей деятельности в значительной степени определяется скоростью трансформации оборотных активов из одного вида в другой. [20]
К настоящему времени получены константы скорости реакции 1-го порядка практически для всех классов органических соединений; значения для некоторых из них приведены в табл. VI-4, а для наиболее изученных - фенолов - в табл. VI-5 и VI-6. Существенно, что эта простая модель ( по которой скорость трансформации вещества линейно зависит от его концентрации) формально согласуется с экспериментальными данными, даже когда скорость трансформации органического вещества в действительности не зависит от его концентрации, а определяется скоростью развития микроорганизмов. Однако это формальное соответствие характерно только для экспоненциальной фазы роста микроорганизмов; в период лаг-фазы и стационарной фазы их развития модель даже чисто формально не соответствует экспериментально наблюдаемой картине. Поэтому при обработке экспериментальных данных соответствующие этим фазам периоды трансформации веществ обычно не учитываются, и количественно оцениваются лишь не имеющие перегиба и плато участк кривых с экспоненциальным снижением концентрации веществ. Однако даже на этих участках кривых модель не адекватна, о чем свидетельствует изменение константы скорости во времени и ее зависимость от начальной концентрации вещества. Следовательно, эта модель и особенно полученные в экспериментах значения константы скорости не могут непосредственно использоваться для расчета скорости самоочищения вод в природных условиях. [21]
Как правило, трансформации происходят между разными штаммами одного и того же вида. Было показано, что в таких случаях скорость трансформации зависит от количества частиц ДНК в трансформирующей среде. Известны также примеры трансформаций между разными видами бактерий, но скорость трансформации в этих случаях в несколько тысяч раз меньше, чем при трансформации внутри одного вида. [22]
Большинство изучаемых в природе термодинамических систем - открытые системы, т.е. способные обмениваться энергией с внешней средой. Классическая термодинамика рассматривает в основном равновесные состояния системы, в которых параметры не изменяются во времени. В открытых же системах реакции и соответствующие энергетические превращения происходят постоянно, поэтому нужно знать скорости трансформации энергии в каждый момент времени. Это значит, что в энергетических расчетах нужно учитывать фактор времени, для чего необходимо сочетать термодинамический и кинетический подходы к описанию свойств открытой системы. Проблема заключается в том, чтобы понять, как связано изменение энтропии с параметрами процессов в открытой системе и выяснить, можно ли предсказать общее направление необратимых процессов в открытой системе по изменению ее энтропии. Главная трудность при решении этой проблемы состоит в том, что необходимо учитывать изменение всех термодинамических величин во времени непосредственно в ходе процессов в открытой системе. [23]
Согласно второму закону термодинамики в изолированной системе энтропия, являющаяся показателем состояния системы и критерием эволюции системы, всегда возрастает. Однако, в природе в большинстве своем системы являются открытыми. В открытых системах может устанавливаться стационарное состояние, при котором необходимо учитывать не только общий статистический баланс энергии, но и скорости трансформации энергии. Это в полной мере относится и к автоколебательным процессам, являющимся самоорганизующимися. Для неустойчивых систем характерна необратимость, повышающая энтропию. В равновесных условиях производство энтропии минимально. Нестабильность возникает из нестабильной динамики. [24]
 |
Изодозные кривые мегавольтного пучка в полиэтилене, нормированные на один электрон, на уровнях 0 1. 0 5 и 1 0 от дозы на поверхности для слаботочного пучка. [25] |
Качественно похожую трансформацию испытывают и энергетические распределения электронного пучка: даже малые толщины вещества заметно уширяют спектр пучка. При толщинах, превышающих половину пробега электронов в графите и 0 15 пробега в меди, распределения приобретают форму, сравнимую с равномерным распределением. При дальнейшем увеличении толщины поглотителя начинают проявляться тенденции относительного увеличения числа низкоэнергетических электронов. Скорость трансформации спектра с ростом глубины проникновения растет по мере увеличения плотности тока пучка. Это, очевидно, объясняется спиралевидным движением электронов в электромагнитном поле, которое увеличивает длину пути и, следовательно, энергетические потери. Большее отличие средней энергии пучка для сильноточного и слаботочного случаев наблюдается в легких материалах. Это объясняется большим сечением рассеяния меди. Меньшее отличие средней энергии сильноточного и слаботочного пучков на больших толщинах связано с возрастанием вклада электронов в приосевой области, где магнитное поле пучка мало. Быстрое размытие спектра электронов сильноточного пучка проявляется также в зависимости вероятной энергии от толщины поглотителя. Вероятная энергия быстрее спадает с толщиной по сравнению со средней энергией вплоть до толщин, выше которых понятие вероятной энергии теряет смысл из-за предельно широкого энергетического спектра. [26]
 |
Элементарные ячейки изотактического полибутена-1. а - форма I, гексагональная. Ъ - форма III. [27] |
При затвердевании из расплава Изотактический полибутен-1 кристаллизуется сначала в тетрагональную структуру, имеющую форму II. Однако со временем в результате старения наблюдается его превращение в форму I, имеющую гексагональное строение. Процесс старения приводит к трансформации начальной тетрагональной структуры спирали 11 / 3 в спирали 3 / 1, что также наблюдается в изотактическом полипропилене. Этот фазовый переход всесторонне изучен [79- 88], и было показано увеличение скорости трансформаций при воздействии механического напряжения и ориентационной вытяжки. [28]
Фенолы поступают в биоценозы также различными путями, в том числе - с промышленными стоками коксохимического производства, некоторых химических производств. Фенолы содержатся в осадках городских сточных вод и могут выщелачиваться оттуда атмосферными осадками, попадая с жидким стоком в почвы и водоемы. Скорость разложения фенолов в почвах довольно велика. Скорость трансформации составляет примерно 40 - 120 мг в сутки. [29]
Приведем теперь кинетические данные Хочкиса и Фокса, полученные с замороженной культурой пневмококков, из которой аликвоты вводились в питательную среду с добавленной трансформирующей ДНК. При этом клетки еще не начинают делиться. Зависимость скорости трансформации от температуры выражается кривой с максимумом при 30 С. [30]
Страницы: 1 2