Скорость - гибель
Cтраница 1
Скорость гибели этих радикалов зависит, по-видимому, от прочности наиболее слабых связей С - Н полимера. Устойчивость концевых радикалов структуры R CH2 - RHC также неодинакова в разных полимерах. [1]
Скорость гибели бактерий в природных реках в значительной степени зависит от гидравлических и биологических характеристик реки. [2]
Скорость гибели макрорадикалов зависит также от их пространственного распределения в объеме полимера и структуры полимерной матрицы. [3]
Скорость гибели гусеницы зависит от степени повреждения кишечника, а также от состава микрофлоры бактерий, размножающихся в теле зараженного насекомого. Гусеницы большой вощинной моли, у которых бактериальная флора более однообразна, погибают после длительного периода развития болезни. [4]
Скорость гибели триплета характеризуется временем полупревращения 1 мкс и не зависит от присутствия мицелл. Однако некоторые растворенные частицы, связанные с мицеллами, способствуют ускорению гибели у В табл. 20.1 приведены данные по переносу энергии триплетного возбуждения на пиренмаслянук кислоту ПМК, пкрен, пирентетрасульфоновую кислоту и ПСК, которые при этом сами переходят в приплетно - озбуж - денное состояние. Скорость переноса триплетной энергии от Ф2 к ПСК зависит от концентрации мицелл при постоянном отношении ПСК / мицелла. [5]
Скорость гибели активных центров, следовательно, зависит от константы скорости роста цепи и вероятности встречи дефекта. [6]
Скорость гибели перекисных радикалов зависит от их структуры. Легче гибнут радикалы, образованные из первичных гидроперекисей. [7]
Если скорость гибели атомов О ( 3Р) измеряется в условиях реакции псевдопервого порядка, когда [ Н2 ] 3 [ О ], то k определяется по уравнению k - [ H2 ] - 4n [ O ] / df и для ее расчета нужно знать только отношения концентраций атомов кислорода. [8]
Поскольку скорость гибели макрорадикалов в твердых полимерах во многих случаях подчиняется закону второго порядка, естественно сделать предположение, что встреча радикалов осуществляется в результате их трансляционной диффузии. [9]
Для скоростей гибели таких полирадикалов имеются две довольно четко разграниченные области температур. В низкотемпературной области ( до 140 - 150 С) энергия активации на - 33 кДж / моль ( 8 ккал / моль) меньше, чем у монорадикалов, а предэкспоненты на 3 - 4 порядка ниже; наблюдаемые константы скорости гибели полирадикалов значительно выше, чем у монорадикалов. В высокотемпературной области константы скорости реакции полирадикалов и монорадикалов сближаются; энергии активации и, предэкспоненты приближаются к соответствующим значениям для монорадикалов. [10]
 |
Изменение относительной концентрации ион-радикалов в зависимости от температуры прогрева ( а и времени освещения ( б. [11] |
Так как скорость гибели в этом случае не зависит от температуры и сохраняется окраска образца, то следует предполагать диссоциацию NH3 в качестве первой ступени гибели в результате передачи энергии экситонами. [12]
При такой скорости гибели лесов более 30 % массивов может исчезнуть уже в начале следующего столетия. [13]
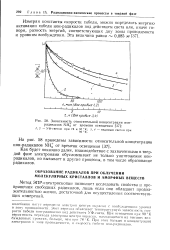 |
Зависимость относительной концентрации ион-радикалов NHJ от времени освещения. [14] |
Измеряя константы скорости гибели, можно определить энергию активации гибели ион-радикалов под действием света или, иначе говоря, разность энергий, соответствующих дну зоны проводимости и уровням возбуждения. [15]
Страницы: 1 2 3 4