Видоспецифичность
Cтраница 1
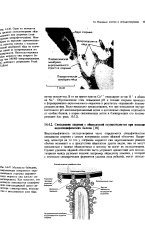
Видоспецифичность оплодотворения часто определяется специфичностью связывания спермия с самым внутренним слоем яйцевой оболочки. Например, несмотря на то что у спермнев морского ежа акросомальная реакция обычно возникает и в присутствии яйцеклеток других видов, они не могут связываться с такими яйцеклетками и поэтому не могут их оплодотворять. [2]
Факторы di не обладают видоспецифичностью, но микроорганизмы разных таксонов различаются как продуктивностью этих ауторегуля-торов, так и чувствительностью к их воздействию. Это позволяет факторам dx выступать в роли ауторегуляторов на уровне микробных сообществ, а также предполагает возможность контролировать развитие промышленных популяций, ассоциаций и консорциумов. [3]
 |
Предполагаемая структура олигосахаридного фактора NodRm-1. Это соединение обусловливает специфический ответ растения-хозяина, в том числе скручивание и деформацию корня. [4] |
К настоящему времени не удалось разработать простых генетических подходов, которые позволяли бы использовать о - гены для повышения конкурентоспособности инокулирую-щих штаммов Rhizobium. Впрочем, можно изменить видоспецифичность бактерий путем переноса nodD - гена из штамма Rhizobium с широкой специфичностью в один из штаммов с узкой специфичностью. Так или иначе, ясно, что образование клубеньков - весьма сложный процесс, и для дальнейшего увеличения конкурентоспособности штаммов Rhizobium потребуются всесторонние исследования с использованием методов генной инженерии. [5]
Из всех имеющихся структурных данных для оксигевиро-ванного и неокснгенированного гемоглобинов можно сделать вывод, что-третичная структура гемоглобина не имеет видоспецифичности. [6]
Физиологическая разнокачественность живых организмов представляет собой фундаментальное условие устойчивого существования жизни как планетарного явления. Теоретически можно представить возникновение жизни в одной форме, но в этом случае запрограммирована конечность жизни как явления: видоспецифичность обмена веществ неизбежно ведет к исчерпанию ресурсов и загрязнению среды продуктами жизнедеятельности, которые невозможно использовать вторично. [7]

Более того, регулируя обмен, можно добиваться заметного повышения выхода целевых продуктов. При этом всегда необходимо учитывать тип дифференцировки, или состояния специализации исходных клеток, так как от него зависит видоспецифичность первичного и вторичного метаболизма. [9]
Интерфероны - клеточные белки и поэтому они видоспеци-фичны, то есть каждому виду животного свойственен свой интерферон, но не являются вирусоспецифическими. При смешанной вирусной инфекции один вирус подавляет другой за счет интер-фероногенности первого - феномен вирусной интефе-р е н ц и и. Иногда эта видоспецифичность очень узкая, например, для курицы, утки, мыщи и крысы, но не перекрестно в группах птиц и грызунов или между группами. Однако есть исключения - человеческий интерферон защищает клетки крупного рогатого скота лучше, чем коровий интерферон. [10]
Оповещение об опасности может быть активным; таковы, в. Специфичность сигналов тревоги подчас обнаруживает их особое биологическое значение. Так, предостерегающий сигнал у обезьян звучит, в отличие от многих других издаваемых ими звуков, резко и кратко. То же свойственно многим другим животным: сигналы тревоги отличаются высокой интенсивностью, но малой длительностью; последнее, очевидно, затрудняет лоцирование сигнала и ориентацию хищника на его источник. Характерно, что при высокой видоспецифичности других звуковых сигналов крики тревоги воспринимаются однозначно многими видами. Это, в частности, хорошо известно для птиц, африканских копытных, пасущихся стадами смешанного видового состава, и для других животных. [11]
Геном млекопитающих содержит несколько разных семейств коротких повторов. Короткие повторы у птиц и амфибий изучены значительно хуже. Число копий коротких повторов, например наиболее изученных повторов Alu-семейства у человека, составляет 3 - Ю5, что соответствует 5 - 6 % массы ДНК клетки. Такие повторы рассеяны по геному и получили название вездесущих. Нук-леотидная последовательность Alu-повтора гомологична последовательности отдельных участков 7S РНК - Структура 7S РНК достаточно консервативна у позвоночных, а гомологии в нуклеотидной последовательности прослеживаются и с 7S РНК насекомых. Поэтому семейства коротких повторов, присутствующие у разных видов, предшественником которых служила 7S РНК, также могут обладать достаточной гомологией. В то же время семейства коротких повторов, как и длинных, характеризуются видоспецифичностью, обусловленной амплификацией той или иной копии клеточных РНК, которые к тому же могли быть по-разному модифицированы в результате процессинга. Локализация ретро-позонов, внедрившихся в отдельные сайты генома у предков млекопитающих, может, по крайней мере, частично сохраняться в процессе дальнейшей эволюции. Например, места локализации Alu-подобного семейства в межгенных промежутках кластера глобиновых генов оказались достаточно сходными у мышей и приматов. [12]
Геном млекопитающих содержит несколько разных семейств коротких повторов. Короткие повторы у птиц и амфибий изучены значительно хуже. Число копий коротких повторов, например наиболее изученных повторов Alu-семейства у человека, составляет 3 - 105, что соответствует 5 - 6 % массы ДНК клетки. Такие повторы рассеяны по геному и получили название вездесущих. Нук-леотидная последовательность Alu-повтора гомологична последовательности отдельных участков 7S РНК - Структура 7S РНК достаточно консервативна у позвоночных, а гомологии в нуклеотидной последовательности прослеживаются и с 7S РНК насекомых. Поэтому семейства коротких повторов, присутствующие у разных видов, предшественником которых служила 7S РНК, также могут обладать достаточной гомологией. В то же время семейства коротких повторов, как и длинных, характеризуются видоспецифичностью, обусловленной амплификацией той или иной копии клеточных РНК, которые к тому же могли быть по-разному модифицированы в результате процессинга. Локализация ретро-позонов, внедрившихся в отдельные сайты генома у предков млекопитающих, может, по крайней мере, частично сохраняться в процессе дальнейшей эволюции. Например, места локализации Alu-подобного семейства в межгенных промежутках кластера глобиновых генов оказались достаточно сходными у мышей и приматов. [13]
Геном млекопитающих содержит несколько разных семейств коротких повторов. Короткие повторы у птиц и амфибий изучены значительно хуже. Число копий коротких повторов, например наиболее изученных повторов Alu-семейства у человека, составляет 3 - 105, что соответствует 5 - 6 % массы ДНК клетки. Такие повторы рассеяны по геному и получили название вездесущих. Нук-леотидная последовательность Alu-повтора гомологична последовательности отдельных участков 7S РНК - Структура 7S РНК достаточно консервативна у позвоночных, а гомологии в нуклеотидной последовательности прослеживаются и с 7S РНК насекомых. Поэтому семейства коротких повторов, присутствующие у разных видов, предшественником которых служила 7S РНК, также могут обладать достаточной гомологией. В то же время семейства коротких повторов, как и длинных, характеризуются видоспецифичностью, обусловленной амплификацией той или иной копии клеточных РНК, которые к тому же могли быть по-разному модифицированы в результате процессинга. Локализация ретро-позонов, внедрившихся в отдельные сайты генома у предков млекопитающих, может, по крайней мере, частично сохраняться в процессе дальнейшей эволюции. Например, места локализации Alu-подобного семейства в межгенных промежутках кластера глобиновых генов оказались достаточно сходными у мышей и приматов. [14]
Страницы: 1