Нуклеотидный состав
Cтраница 1
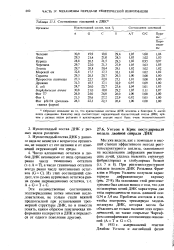
Нуклеотидный состав различных компонент рРНК неодинаков и не подчиняется правилам Чаргаффа. По-видимому, вторичная структура рРНК характеризуется многочисленными короткими двуспиральными участками, соединенными - промежуточными, однотяжевыми участками в гибкую палочку. Молекулы белка взаимодействуют с неспиральными участками рРНК, не нарушая ее вторичной структуры, и образующийся таким образом рибонуклеопротеидный комплекс сворачивается в компактную структуру рибосомной субъединицы. В 705-рибосоме содержится около 65 полипептидных цепей со средним молекулярным весом каждой цепи 65000, в SOS-рибосоме число таких цепей вдвое больше. Установлена частично глобулярная структура этих белков и наличие в них а-спиральных участков. [1]
Нуклеотидный состав ДНК, выделенных из организмов разных видов, сильно различается, но является характерным для каждого вида. Видоспецифич-ность ДНК - основа геносистематики и используется для установления филоге-нетич. [2]
Нуклеотидный состав РНК обычно определяют после щелочного гидролиза препарата, дающего смесь свободных рибонуклеотидов. [3]
Нуклеотидный состав ДНК может быть определен спектрофотометриче-ским методом. [4]
 |
Соотношение оснований в ДНК1. [5] |
Нуклеотидный состав ДНК у данного вида не меняется с возрастом организма, не зависит от его питания и от изменений окружающей его среды. [6]
 |
Двойная спираль ДНК ( стрелками показано направление полинук-леотндной цепи. [7] |
Нуклеотидный состав ДНК подчиняется ряду правил ( т.наз. правила Чаргаф-фа), важнейшее среди к-рых - одинаковое содержание А и Т, О и С у любой клеточной ДНК. Нуклеотидный состав РНК подобным правилам не подчиняется. [8]
Нуклеотидный состав ДНК микроорганизмов дает возможность решить вопрос о принадлежности микроорганизмов к тому или иному виду. [9]
 |
Цепь РНК. [10] |
Нуклеотидный состав нуклеиновой кислоты устанавливается путем гидролиза. [11]
Поскольку нуклеотидный состав рибосомной РНК не коррелирует с нуклеотидным составом ДНК в тех же клетках, казалось сомнительным, чтобы и рибосомная РНК действительно образовывалась на ДНК-матрице. Яновский и Спигелман выбрали организмы, ДНК которых резко отличается по составу от рибосомной РНК, пометили их РНК с помощью Р32 или Н3 и позволили затем клеткам находиться на нерадиоактивной среде еще некоторое время. За этот период метка исчезала из быстро метящейся m - РНК и накапливалась в стабильной r - РНК. Затем дали возможность образоваться гибридам, после чего удалили РНК-азой всю негибридизированную РНК и выделили устойчивые к РНК-азе гибриды. Оказалось, что только менее 0 4 % всей ДНК комплементарно РНК из 16S - и 238-рибосом. Нерибосомная РНК из данного организма не конкурирует за эту фракцию ДНК. Более того, сами r - РНК из 16S - и 238-рибосом берут начало от различных участков ДНК. Это предположение основано на следующих фактах: во-первых, максимальное количество РНК, способное образовать гибрид с ДНК, различно для этих двух РНК. [12]
Какой нуклеотидный состав будет иметь первая вновь синтезированная комплементарная цепь и цепь синтезированная после неограниченного числа репликаций. [13]
Однако нуклеотидный состав ДНК - признак в основном крупномасштабный, кроме того, могут встречаться случайные совпадения состава ДНК у далеко отстоящих родов, поэтому большое внимание в монографии уделяется методу молекулярной гибридизации ДНК применительно к целям систематики. [14]
В скобках указан нуклеотидный состав ДНК из того ше источника. [15]
Страницы: 1 2 3 4