Исходное состояние - реакция
Cтраница 1
Исходное состояние реакции - комплекс рутения ( лиганды не обозначены), молекула Н2 и молекула фумаровой кислоты. [1]
В активных центрах ферментов в рамках относительно жесткой третичной структуры белка взаимодействующие функциональные группы уже в исходном состоянии реакции в гораздо большей степени сближены и сориентированы, чем в большинстве неферментативных внутримолекулярных процессов. [2]
Умозрительно вклад концентрационного и ориентационного эффектов в ускорение внутримолекулярной реакции можно было бы отделить от того вклада, который обусловлен частичным преодолением энергетического барьера реагентами уже в исходном состоянии реакции. Следует полагать, что первый вклад приводит к более благоприятной энтропии активации, а второй обусловливает понижение энтальпии активации внутримолекулярной реакции. [4]
 |
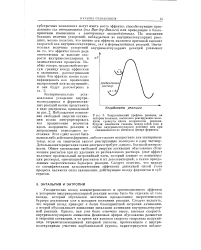
Диаграмма изменений стандартной свободной энергии реакции ( 1 - й путь и ( 2 - й путь с учетом эффекта сближения и ориентации реагентов ( ср. с 12. [5] |
Казалось бы в таком случае, что эффективность ферментативного катализа должна возрастать при увеличении потенциальной свободной энергии внутримолекулярного взаимодействия ( - ДО внут Р) Это действительно происходит в ферментативных реакциях второго порядка ( см. § 2 этой главы), однако при [ RY ] KS, когда исходное состояние реакции - - это фермент-субстратный комплекс, такое требование не достаточно. [6]
Этот своеобразный компенсационный эффект открывает, казалось бы, довольно простой путь к созданию высокоактивных катализаторов: достаточно синтетическим путем объединить в одну молекулу хотя бы две функциональные группы, потенциально способные одновременно [ по механизмам типа (3.5) или (3.6) 1 участвовать в образовании переходного состояния. Включение их в одну молекулу уже в исходном состоянии реакции может в значительной степени снизить энтропийные потери, необходимые при построении переходного состояния. [7]

Кроме того, нельзя считать правомерным использовавшееся при выводе всех предыдущих уравнений до-лущение о постоянстве AG в одной реакционной серии. При замене субстрата это допущение скорее всего нарушится, поскольку изменение в исходном состоянии реакции будет неизбежно сказываться на ее скорости. [9]
Согласно оценке, проведенной Ласковским и Шерага [72], изменение энтропии на каждую замороженную связь в боковой группе аминокислотного остатка, связанного с жесткой полипептидной цепью, составляет примерно - ( 4 - 7) кал / моль / град [ - ( 16 8 - 29 4) ] Дж / моль / град. Поэтому исходя из найденной величины ДД5з - 10 - 12 кал / моль / град ( 42 - 50 4 Дж / моль / град) можно полагать, что в случае специфических субстратов, содержащих а-ацетиламидную группу, происходит уже в исходном состоянии реакции ( в ацилферменте или, соответственно, в комплексе Михаэлиса) почти полное, благоприятное для реакции замораживание всего фрагмента - С-С ( О) - О -, центральная карбонильная группа которого претерпевает химические превращение. [10]
Из уравнения (2.21) видно, что термодинамически эффективность ферментативного катализа определяется разницей свободных энергий межмолекулярного ( при образовании комплекса Михаэлиса) и внутримолекулярного ( в переходном состоянии реакции) образования связи E-R. Это связано с тем ( см. рис. 12), что барьер свободной энергии активации ферментативной реакции ( Л0 внутр) в этом случае уменьшается ( по сравнению с Д ( 5п) и, следовательно, скорость процесса [ уравнение (2.20) ] возрастает. Наоборот, при заданном значении AG BHyTP термодинамически более благоприятное взаимодействиеЕ - R в исходном состоянии реакции ( фермент-субстратный комплекс XE-RY) будет тормозить ее протекание. Это связано с тем, что активационный барьер AGi BnyTP ( см. рис. 12), определяющий скорость превращения фермент-субстратного комплекса [ уравнение (2.20) ], при этом возрастает. [11]
Перечисленные физические условия показывают, что отнюдь не любая система способна к производству, хранению и переработке информации. Например, сразу же отпадают обширные классы консервативных гамильтоновых систем. К системам, удовлетворяющим условиям 1 - 4, относятся переключательные схемы электроники и микроэлектроники, а также мультистабильные реакции. Идеальными информационными системами естественной эволюции являются реакции синтеза биополимеров. Биомолекулы представляют собой гетерополимеры с длиной цепи от 102 до 106 элементарных звеньев. Нейтральное исходное состояние реакции синтеза представляет собой смесь свободных элементарных звеньев необходимого брутто-состава. Состояния-аттракторы этой информационной системы соответствуют химически устойчивым конфигурациям биополимеров. [12]
Страницы: 1