Высокая специфичность - фермент
Cтраница 1
Высокая специфичность фермента определяется его белковой частью. Коферменты значительно менее специфичны и поэтому могут катализировать различные реакции в зависимости от того, с каким апоферментом они связаны. [1]
Высокая специфичность фермента определяется его белковой частью. Коферменты значительно менее специфичны, и поэтому они могут катализировать реакции в зависимости от того, с каким апо-ферментом они связаны. Число коферментов по сравнению с ферментами невелико, однако они отличаются разнообразным химическим строением. [2]
 |
Схема адсорбции субстра - - Адсорбционные. [3] |
Как уже сказано, высокая специфичность ферментов проявляется не только по отношению к химическому составу реагентов, но и к их пространственному строению. Например, если молекула продукта реакции содержит асимметрический атом, а молекула исходного субстрата симметрична, значит фермент осуществляет асимметрический синтез одного изомера. Если же исходное вещество представляет собой рацемическую смесь, фермент ускоряет превращение только одного изомера. Если в качестве исходных веществ берут цис - и гранс-изомеры, высокоспецифичный фермент изменяет скорость реакции только одного стереоизомера. [4]
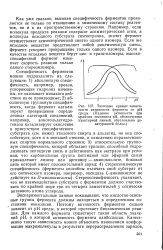 |
Типичные кривые зависимости активности фермента от рН среды. ( Падение активности при крайних значениях рН, обозначенное пунктирной линией, обусловлено денатурацией белка. [5] |
Как уже сказано, высокая специфичность ферментов проявляется не только по отношению к химическому составу реагентов, но и к их пространственному строению. Например, если молекула продукта реакции содержит асимметрический атом, а молекула исходного субстрата симметрична, значит фермент осуществляет асимметрический синтез одного изомера. Если же исходное вещество представляет собой рацемическую смесь, фермент ускоряет превращение только одного изомера. Если в качестве исходных веществ берут цис - и транс-изомеры, высокоспецифичный фермент изменяет скорость реакции только одного стереоизомера. [6]
Во-вторых, на этом примере видны структурные основы высокой специфичности ферментов, в частности стереоспецифичнбсти. [7]
Мультиплетность ( полифункциональность) ферментативного катализа и есть, по-видимому, причина высокой специфичности ферментов. Если для осуществления каталитической реакции обязательным требованием является образование нескольких химических связей между определенными атомами молекулы субстрата и активного центра фермента, то отсюда вытекает требование строгой комплементарности реагирующих молекул. [8]
В клетке, содержащей сотни метастабильных соединений, должно было бы происходить бесконечно большое число реакций, однако число это ограничено в силу высокой специфичности ферментов. Таким образом, присутствующие ферменты полностью определяют характер реакций, протекающих в живой клетке. Состав биологической системы, несомненно, определяется специфичностью, концентрацией и активностью ее ферментов. Изменения специфичности фермента возможны только в результате мутаций. [9]
Все разнообразные обратимые окислительно-восстановительные реакции субстратов осуществляются почти исключительно при участии двух кофакторов - НАД и НАДФ. Высокая специфичность фермента к субстрату обусловливается почти исключительно структурой белка - последовательностью соединения L-аминокислот и их пространственного расположения. Молекулярная масса ферментов составляет от 84 000 для алкргольдегидрогеназы из печени до 1 000 000 для глутаматдегидрогеназы из митохондрий печени быка. [10]
В катаболических и анаболических превращениях одна реакция следует за другой в строжайшей последовательности, так как продукты реакции предыдущей стадии процесса, как правило, являются субстратом для последующей. Такая четкая преемственность возможна благодаря высокой специфичности ферментов, участвующих в обмене веществ. [11]
Однако эта интересная теория, несмотря на большое количество работ, проведенных на протяжении последних 15 лет, не способствовала изысканию новых полезных химиотерапевтических средств, в то время как многие вещества, синтезированные без теоретических предпосылок, оказались антиметаболитами. В некоторых случаях неудачи могли быть обусловлены высокой специфичностью ферментов в использовании незаменимых метаболитов. В других случаях недостаточная химиотераповтическан активность вещества in vivo могла быть следствием избытка незаменимых метаболитов в животном организме. Например, сульфокпслота ( XXII), аналог нантотеновой кислоты, применяемая как антиметаболит пантотеновой кислоты, подавляет рост гемолитического стрептококка in ri / ro и предохраняет от гибели крыс, зараженных этим же видом микроба, но не защищает от смерти мышей, так как количество пантотеновой кислоты в крови мышей выше, чем в крови крыс. Другая трудность, возникающая при применении теории антиметаболитов как основы изыскания химиотерапевтических веществ, заключается в том, что многие незаменимые метаболиты микробов являются в то же время витаминами в питании человека. Антиметаболнты в этом случае могут вызвать симптомы недостатка витаминов в организме хозяина и не будут поэтому обладать избирательной токсичностью. [12]
Ферменты обладают высокой специфичностью действия. Это свойство часто существенно отличает их от неорганических катализаторов. Так, мелкоизмельченные платина и палладий могут катализировать восстановление ( с участием молекулярного водорода) десятков тысяч химических соединений различной структуры. Высокая специфичность ферментов обусловлена, как было отмечено, конфор-мационной и электростатической комплементарностью между молекулами субстрата и фермента и уникальной структурной организацией активного центра, обеспечивающими узнавание, высокое сродство и избирательность протекания одной какой-либо реакции из тысячи других химических реакций, осуществляющихся одновременно в живых клетках. [13]
Оказалось, что цепные механизмы реакции не играют существенной роли в биологическом катализе. После установления химической природы ферментов подтвердилось представление, выдвинутое более 80 лет назад В. Благодаря высокому сродству связывания и образованию ES-комплекса резко возрастает число молекул субстрата, вступающих в реакции. Фишера, которую иногда называют теорией жесткой матрицы. Таким образом, жесткая структура активного центра оказывается комплементарной молекулярной структуре субстрата, обеспечивая тем самым высокую специфичность фермента. [14]
Гипотеза аллостерического торможения, вы -) двинутая Mono, Шанже и Жакобом [10], хорошо объясняет поведение описанных выше и других - ферментных систем. Согласно этой гипотезе, цэбщей моделью аллостерического белка служит гипотетическая ферментная молекула, обладающая по крайней мере двумя стереоспецифи-чески различными, неперекрывающимися рецеп-торными центрами. Он связывает субстрат и определяет биологическую активность белка. Другой, алло-стерический, центр специфически и обратимо присоединяет аллостерический эффектор. Алло-стерический эффектор не обязательно должен иметь химическое или метаболическое родство с субстратом фермента. Предполагается, что эффектор вызывает изменение конформации активного центра, не соединяясь с ним и не изменяя специфичности этого центра. Теория индуцированного соответствия, предложенная Кошландом, объясняет высокую специфичность фермента по отношению к субстрату. [15]
Страницы: 1