Спорозоит
Cтраница 3
После достижения определенного размера трофозоиты отделяются от эпителия, округляются, и происходит деление их. Ядра мигрируют к поверхности трофозоита, который распадается на большое количество банановидных меронтов. Меронты вновь оседают на эпителий и вырастают в булавовидные трофозоиты. Последние округляются, соединяются друг с другом и образуют шаровидные или сферически-овальные цисты. Внутри цист образуются пузыревидные споры, в которых расположены спорозоиты, соединенные задними концами со сферическим остаточным телом. Яйцевидно-удлиненные, с тонкой оболочкой, споры не имеют полярного отверстия и открываются, по-видимому, при разрыве оболочки. Из комаров выделено много видов этих паразитов. Вид Caulleryella anophelis Hesse с 8 - 11 спорами в цисте найден в Anopheles claviger Meig. [31]
Когда тело меронта достаточно нарастает, оно округляется, и возникает сферический гаметоцит. Гаметоциты соединяются в пары таким образом, что плоскости их соприкосновения образуют экватор удлиненной цисты. В цисте гаметоциты остаются отделенными друг от друга, пока происходит деление ядер на два. Деление редукционное, и после слияния двух гаплоидных гамет образуется шаровидная зигота, которая позднее удлиняется в веретеновидную, приобретая форму будущей споры. Ядро в ней делится на 8 дочерних ядер и образуется 8 червеобразных спорозоитов. Циста не имеет прочной оболочки. Оболочка споры утолщенная, на полюсах имеются чечевицеобраз-ные утолщения. У краев этой чечевицы оболочка споры тоньше, и здесь она разрывается при вскрытии споры под воздействием желудочного сока хозяина. [32]
Из сферических 4 - 6-ядерных схизонтов развиваются гамето-циты и соединяются попарно, причем один гаметоцит всегда серповидно обвивает другого. Здесь же можно обнаружить случаи, когда один гаметоцит ( мужской) образует оболочку другого. Возникающая шаровидная циста отличается большим количеством хорошо заметных ядер, вокруг которых постепенно образуется 8 - 16 или 32 споры, объединенные тонкой пленкой в шаровидную цисту. Эта пленка при изготовлении мазков разрывается, и в препарате всегда видны только одиночные споры веретеновидной формы, размером 13X6 мк, закрытые на полюсах шаровидными про-бочками, подобно яйцам власоглава. Споры открываются в этих пробочках, и из них выходит 8 булавовидных спорозоитов. Название Triboliocystis garnhami Dissanaike является синонимом Farinocystis tribolii Weiser. Заражение паразитом происходит в 1 - 4 - м возрасте личинок, причем заражение в первых трех возрастах приводит к гибели личинок перед их последней линькой, заражение в последнем возрасте наблюдалось лишь в единичных случаях. Распространение болезни нарастает до последнего возраста личинок и к их 5-му возрасту бывает заражено более 90 % особей. При искусственном разведении болезнью поражается 2 - 5 % взрослых жуков. Такое развитие болезни объясняется пищевыми связями в популяции. Личинки жуков Tribolium поедают все белковые и жировые примеси в муке или зерне для пополнения кормового рациона этими дефицитными веществами. Они поедают мертвых личинок своего вида, а также остатки и трупы насекомых и других видов, погибших от этой болезни и заполненных спорами схизо-грегарины, и таким путем заражаются. Степень распространения болезни в популяции повышается с увеличением количества погибших от нее особей, а частота встречаемости спор схизогрега-рины в муке является фактором второстепенного значения. [33]
В кишечник мухи поступает кровь с эритроцитами, зараженными гаметоцитами гемоспоридий. Почковидный макрогаметоцит освобождается, а микрогаметоцит делится на несколько тонких червеобразных микрогамет, которые стремятся найти макрогаметы. Возникающая при соединении микро - и макрогаметы зигота, вытягиваясь, принимает яйцевидную форму, затем образует оокинету, которая проходит через эпителий кишечника и проникает в. Внутри ооцисты происходит многократное деление ядра, и плазматическая масса делится на большое количество спорозоитов, собранных в правильные пучки, расположенные по всей цисте. Через несколько дней оболочка цисты лопается, и спорозоиты мигрируют по телу хозяина, достигая задней части слюнных желез мухи, расположенных в грудных и первом брюшном сегментах. При высасывании мухой крови позвоночного животного спорозоиты вместе со слюной насекомого попадают в ранку в коже позвоночного хозяина и заражают его. Развитие гемоспоридий в насекомом продолжается несколько недель, спорозоиты в слюнных железах сохраняют жизнеспособность в течение всей зимовки насекомого. [34]
В кишечник мухи поступает кровь с эритроцитами, зараженными гаметоцитами гемоспоридий. Почковидный макрогаметоцит освобождается, а микрогаметоцит делится на несколько тонких червеобразных микрогамет, которые стремятся найти макрогаметы. Возникающая при соединении микро - и макрогаметы зигота, вытягиваясь, принимает яйцевидную форму, затем образует оокинету, которая проходит через эпителий кишечника и проникает в. Внутри ооцисты происходит многократное деление ядра, и плазматическая масса делится на большое количество спорозоитов, собранных в правильные пучки, расположенные по всей цисте. Через несколько дней оболочка цисты лопается, и спорозоиты мигрируют по телу хозяина, достигая задней части слюнных желез мухи, расположенных в грудных и первом брюшном сегментах. При высасывании мухой крови позвоночного животного спорозоиты вместе со слюной насекомого попадают в ранку в коже позвоночного хозяина и заражают его. Развитие гемоспоридий в насекомом продолжается несколько недель, спорозоиты в слюнных железах сохраняют жизнеспособность в течение всей зимовки насекомого. [35]
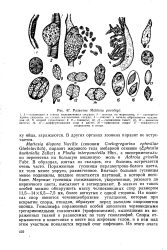 |
Развитие Mattesia povolnyi. [36] |
В образцах, взятых из складов, эта болезнь встречается очень часто. Пораженные гусеницы перламутрово-белого цвета, их тело менее упруго, размягченное. Вначале больные гусеницы менее подвижны, позднее оплетаются паутиной, в которой погибают. Мертвые гусеницы съежившиеся, укороченные, розового до кирпичного цвета, высыхают и затвердевают. На полюсах спор имеются чечевицеобразные утолщения, которые при вскрытии споры, отпадая, образуют перед выходом спорозоитов оконца. Гемолимфа пораженных особей заполнена червовидными грегариноидными схизонтами ( трофозоитами), вышедшими из пораженных тканей и разносимых по телу гемолимфой. Споры открываются в кишечнике в месте под жировым телом, и в нем над этим участком появляется первый очаг инфекции. [37]
В более старых цистах можно уже обнаружить восковидную структуру, разделяющую плазмодии, которые образуются из первоначальных веретеновидных или яйцевидных отрезков в виде шаровидных образований, содержащих максимум 8 ядер. В этой фазе ядра уже более мелкие, компактные иногда удлиненные. Поверхностная оболочка этих плазмодиев уплотняется, содержимое хуже окрашивается, затем образуются вначале чечевицевид-ные, позднее приплюснутые с одной стороны споры ( собственно панспоробласты или цисты) в плазме, которые содержат по 8 ядер. Оболочка этих спор трудно проницаема для красителей. Размеры спор 5 - 7X4 мк, содержимое спор плохо различимо под микроскопом, при фиксации споры деформируются. В литературе отсутствуют данные о структуре спор и о происходящих в них изменениях; можно предполагать, что внутри спор образуется 8 одноядерных спорозоитов. Перегородки, образовавшиеся при споруляции между панспоробластами, постепенно автолизируют-ся, оболочка пораженной эпителиальной клетки разрывается, и высвободившиеся из клетки споры поступают в полость кишечника, а затем с экскрементами выделяются наружу. О патологическом действии возбудителя на зараженных жуков пока ничего неизвестно. [38]
Спорогония начинается с того, что в цисте после слияния гамет вокруг каждой зиготы образуется тонкая пленка, которая постепенно затвердевает; из нее формируется оболочка споры. Внутри споры одноядерный споробласт вновь делится, и возникают 4 - 8 червеобразных спорозоитов с ядром, лежащим в первой трети их длины. У спор некоторых видов спорозоиты, расположенные по длине споры, укреплены на двух ножках, выходящих из вершин споры. У других видов все спорозоиты подвешены тонким концом к одному полюсу остаточного тела споры и висят, подобно грозди бананов. Форма споры соответствует расположению в ней спорозоитов. Веретеновидные споры содержат спорозоиты, расположенные в чередующемся порядке, сидящие на ножках, иногда слегка спирально скрученные. Если спорозоиты собраны в гроздья, заключающие их споры широкояйцевидной формы, с тонкой оболочкой, разрывающейся при вскрытии споры. [39]
Род Diplocystis Kunstler характерен тем, что трофозоиты уже в ранней стадии округляются и образуют цисты из двух сферических, расположенных рядом индивидов. Споры почти шаровидные, с восемью спорозоитами, открываются в кишечнике хозяина, освобождающиеся спорозоиты проникают в стенки кишечника и выседают на базальной мембране. Здесь они соприкасаются, соединяются попарно в сизигии и образуют цисты. Весь период развития циста состоит из двух шариков, которые образуют отдельные зигонты. Цисты увеличиваются в размере и выдаются из эпителия в виде выростов диаметром до 2 мм, затем выпадают из эпителия кишечника в его полость. Здесь они дозревают и образуются гаметы, которые после многократного деления ядер соединяются и начинается редукционное деление. Споры образуются внутри цисты: у Diplocystis schnei-deri Kunst, они широкояйцевидные, длиной 7 - 8 мк, с продольным швом в форме буквы Y. Высвободившиеся спорозоиты достигают 16 мк. Паразит развивается в тараканах Periptaneta americana L. Cuenot живет в домовых сверчках. [40]
Этот род был описан по виду, который поражает водных клещей из рода Hydrachna. Menzbieria chalcographi Weiser известна из жирового тела короеда Pityogenes chalcographus L. Формы, развивающиеся при схизогонии, очень близки формам рода Mat-tesia. Сферические многоядерные схизонты образуют рой червеобразных меронтов, проникающих в окружающие жировые ткани и вырастающих в новые плазмодии с пенистой плазмой и крупными ядрами. После второй схизогонии меронты округляются и вырастают в сферические гаметы, которые соединяются в шаровидных цистах. В цисте образуется 62 споробласта, лежащих на поверхности зиготы, из остатков которой в середине формируется остаточное тело. На одном полюсе имеется небольшая пора, через которую выходят спо-розоиты. В одной споре содержится 8 червеобразных спорозоитов длиной 12 - 15 мк. [41]
В кишечник мухи поступает кровь с эритроцитами, зараженными гаметоцитами гемоспоридий. Почковидный макрогаметоцит освобождается, а микрогаметоцит делится на несколько тонких червеобразных микрогамет, которые стремятся найти макрогаметы. Возникающая при соединении микро - и макрогаметы зигота, вытягиваясь, принимает яйцевидную форму, затем образует оокинету, которая проходит через эпителий кишечника и проникает в. Внутри ооцисты происходит многократное деление ядра, и плазматическая масса делится на большое количество спорозоитов, собранных в правильные пучки, расположенные по всей цисте. Через несколько дней оболочка цисты лопается, и спорозоиты мигрируют по телу хозяина, достигая задней части слюнных желез мухи, расположенных в грудных и первом брюшном сегментах. При высасывании мухой крови позвоночного животного спорозоиты вместе со слюной насекомого попадают в ранку в коже позвоночного хозяина и заражают его. Развитие гемоспоридий в насекомом продолжается несколько недель, спорозоиты в слюнных железах сохраняют жизнеспособность в течение всей зимовки насекомого. [42]
Червеобразные схизонты разрастаются в грушевидные, позднее в широкояйцевидные схизонты, ядра которых делятся, образуя большое количество дочерних ядер. Вокруг этих ядер формируются и дифференцируются сигаровидные мерозоиты с ядром посредине. Оболочка схизонта лопается, и освободившиеся мерозоиты переходят в ткани, по которым они медленно расселяются вокруг. Во вновь пораженных клетках мерозоиты утолщаются, приобретая ладьевидную, яйцевидную, а затем шаровидную форму. Деление вновь повторяется в зависимости от пространства, которым располагает паразит в жировом теле хозяина. При последующем делении возникают мерозоиты, которые, вырастая, преобразуются в половые стадии, шаровидные гаметоциты. Часть из них остается неизменными, а лишь редуцирует хроматин ядра и таким способом образует гаплоидные макрогаметы, ядро других делится на 64 - 128 дочерних ядер. Макрогамету оплодотворяет только одна микрогамета, хотя в некоторых случаях на макрогамету стремятся осесть несколько микрогамет. После разделения шаровидных споробластов в центре цисты ( зиготы) остается бесформенное остаточное тело. В каждой споре зародыш делится и образуются два спорозоита, изогнутые кольцом вокруг шаровидного остаточного тела. [43]
Плазма мужских особей ацидофильна, и они окрашиваются красителем Гимза в красноватый цвет, а женские особи в синеватый. Обе особи бывают соединены последовательно друг за другом, рядом, задними частями тела или различными отростками. Их тела укорачиваются и расширяются, создавая общее шаровидное образование, покрывающееся прочной оболочкой, и возникает циста. Внутри цисты оба индивида изменяются. Их ядра делятся на большое количество дочерних ядер, масса плазмы перегруппировывается, ядра перемещаются к поверхности извилисто разделенной массы гамонтов. Вокруг последних плазма концентрируется в шаровидные или удлиненные сгустки. В одних сгустках образуются мужские, в других женские гаметы, которые стремятся друг к другу и сливаются. В центре остается относительно большое остаточное тело с остатками раздувшихся ядер. Зиготы, возникшие от слияния гамет, округляются и покрываются оболочкой, которая постепенно затвердевает и приобретает форму созревшей споры. Дозревание цисты и находящихся в ней спор ( спороцисты) происходит обычно вне тела хозяина, в его экскрементах. У ряда видов споры выходят из хозяина еще как одноядерные споробласты и дозревают лишь при доступе воздуха, образуя внутри спорозоиты. [44]
Страницы: 1 2 3