Сращивание - цепь
Cтраница 2
Сращивание цепей допускается путем электро - и кузнечно-горновой сварки новых вставленных звеньев или с помощью специальных соединительных звеньев. После сращивания цепь должна быть испытана. [16]
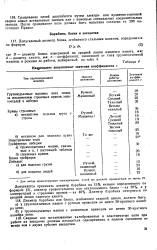 |
Наименьшие допускаемые значения коэффициента е. [17] |
Сращивание цепей допускается путем электро - или кузнечно-горновой сварки новых вставленных звеньев или с помощью специальных соединительных звеньев. После сращивания цепь должна быть испытана согласно ст. 290 настоящих Правил. [18]
Обычно сращивание цепей осуществляется посредством атомов серы или кислорода, которые способны присоединяться одновременно к двум цепям, образуя между ними своеобразные мостики. [19]
Сращивать грузовые канаты запрещается. Сращивание цепей ( гри разрыве или замене отдельных звеньев) допускается при помощи кузнечной сварки или специальных соединительных звеньев. [20]
На заводах-изготовителях цепи испытывают под нагрузкой, равной 50 % их разрывной нагрузки. Износ звена цепи допускается не более 10 % первоначального диаметра. Цепи с вытянутыми звеньями или трещинами применять запрещается. Сращивание цепей возможно вставкой новых или специальных соединительных звеньев, но после этого цепь вновь испытывают. [21]
Цепные стропы по сравнению с канатами более долговечны. Изнашивание звена сварной или штампованной цепи допускается не более 10 % от первоначального диаметра. После сращивания цепи испытывают статической нагрузкой, в 1 25 раза превышающей их номинальную грузоподъемность. [22]
Мак Клинток, которая в опытах с кукурузой показала, что гены могут перемещаться в пределах генома и влиять на механизмы экспрессии. В дальнейшем было установлено, что это явление характерно для многих эукариотических и прокариотических клеток. Транспозоны эукариотических клеток гораздо больше и включают в себя набор различных генов. Внутригеномное перемещение и встраивание транспозонов требует разрыва и последующего сращивания цепи ДНК. Репликация транспозона в одном сайте цепи, а затем перемещение и репликация в другом создают благоприятные возможности для дальнейших гомологичных рекомбинаций в клетке. [23]
Для объяснения указанных явлений плодотворны механо-химй-ческие представления, рассматривающие глинистые агрегаты как блоки макромолекул. Их анизометрия и микродефекты обусловливают неравномерное распределение напряжений даже при весьма малых деформациях. На отдельных участках они значительно превышают молекулярные силы, скрепляющие между собой агрегаты и пачки частиц, и могут даже достигать критических значений, больших, чем энергия ковалентных связей, действующих внутри решетки. Это приводит к разрыву агрегатов. И здесь деструкция идет лишь до определенного предела с выдедением объемных фрагментов, величина которых определяется числом кристаллохими-ческих дефектов. При растяжении или сдвиге внутри пачек в первую очередь нарушаются связи между отдельными блоками, но по мере возрастания межатомных расстояний происходит разрыв ковалентных связей, что вызывает механическую активизацию химических реакций. Например, А. С. Кузьминский установил, что при окислении растянутого каучука энергия активации падает до 3 ккал / моль. В результате становятся возможны реакции, типичные для свободных радикалов. У глины это может усилить ее реакционную способность. У классических полимеров при отсутствии акцепторов наиболее вероятны рекомбинации, сращивание цепей, восстановление ковалентных связей. [24]
Страницы: 1 2