Температурная адаптация
Cтраница 1
Температурная адаптация - служит для компенсации ошибки регулирования частоты вращения или момента двигателя в замкнутой системе управления, возникающей при температурных изменениях сопротивлений обмоток статора и ротора. При отсутствии встроенного датчика температуры обмоток статора расчет сопротивлений осуществляется по заложенным в преобразователь тепловой и электрической моделям двигателя. [1]

На фоне температурных адаптации общего типа, настраивающих метаболические системы на существование в определенных режимах окружающей температуры, у многих пойкилотермных организмов ( особенно у животных) функционируют специализированные адаптивные реакции, лабильно отвечающие на относительно быстрые и кратковременные изменения внешней температуры. В частности, это относится к использованию эндогенного теплообразования для повышения и некоторой стабилизации ( хотя бы временной) температуры тела Многие виды пойкилотермных животных используют тепло, образующееся при работе локомоторной мускулатуры, для создания временной независимости температуры тела, а соответственно и уровня метаболизма от колебаний температуры среды. [3]
У пойкилотермных животных стабильный тип температурной адаптации представлен в виде настройки температурного оптимума деятельности ферментов и уровня теплоустойчивости тканей. На этом фоне сохраняется эффективность функциональной реакции на действие конкретных температур. [4]
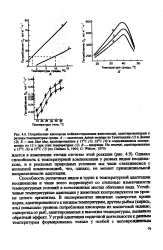
Способность различных видов и групп к температурной адаптации неодинакова и чаще всего коррелирует со степенью изменчивости температурных условий в естественных местах обитания вида. Устойчивые температурные адаптации у животных контролируются на уровне целого организма. [6]
Относительно преимуществ гетерозиготности по белкам с точки зрения температурной адаптации была выдвинута гипотеза, согласно которой организмы, живущие в условиях непредсказуемого температурного режима, будут проявлять тенденцию к большей генетической изменчивости - у них будет больше ал-лоферментов, чем у организмов, живущих в стабильной среде. Эта гипотеза основана на предположении, что различные алло-ферменты данного фермента будут особенно хорошо работать в разных диапазонах температуры. Поэтому мы могли бы ожидать, что, подобно тому как у форели есть два генных локуса для теплового и холодового вариантов ацетилхолинэстеразы, найдутся и такие случаи, когда один аллель определенного гена кодирует тепловой аллофермент, а другой аллель - холодо-вой аллофермент. [7]
Как и следовало ожидать исходя из аналогии с температурной адаптацией, эта рыба обладает ферментами, позволяющими ей переносить большие различия в давлении. Независимость от давления распространяется здесь как на свойства, связанные с числом оборотов, так и на сродство к лигандам ( табл. 23), но существует, по-видимому, известный предел возможного снижения таких параметров, как А У и энергия активации. Таким образом, важнейшие пути адаптации ферментов к давлению, так же как и к температуре, связаны главным образом с изменением взаимодействий между ферментом и лигандами. [8]
Изучение нуклеотидного состава нуклеиновых кислот у различных организмов показало, что, несмотря на значительную изменчивость содержания Г - Ц, не существует сильно выраженной корреляции между процентом Г - Ц в ДНК и температурной адаптацией. Однако в случае рибосомной РНК ( рРНК) дело обстоит иначе: по крайней мере у термофильных бактерий, по-видимому, происходил отбор на высокое относительное содержание гуанина и цитозина. Это различие между ДНК и рРНК, возможно, с язано с тем, что молекула ДНК образует двойную спираль на всем своем протяжении, тогда как рРНК имеет более ограниченную вторичную структуру - основания в ней спарены лишь на коротких участках цепи. Поскольку целостность систем, стабилизируемых слабыми связями, зависит от всей совокупности таких связей, возможно, что каждая отдельная связь, участвующая в поддержании вторичной структуры рРНК, имеет относительно большее значение, чем такая же связь в ДНК. [9]
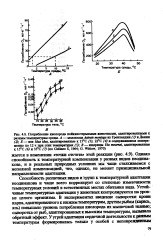
Способность различных видов и групп к температурной адаптации неодинакова и чаще всего коррелирует со степенью изменчивости температурных условий в естественных местах обитания вида. Устойчивые температурные адаптации у животных контролируются на уровне целого организма. [11]
Принципиально иной тип приспособления к температурному фактору свойствен гомойотермным животным. У них температурные адаптации связаны с активным поддержанием постоянства внутренней температуры и основаны на высоком уровне метаболизма и эффективной регулирующей функции центральной нервной системы. [12]
В эксперименте способность пойкилотермных животных к выбору оптимальных температур отчетливо демонстрируется в приборах, где создается градиент температур. Опыты с рыбами, рептилиями, насекомыми и другими животными показывают специфичность предпочитаемых температур для отдельных видов и популяций. При этом отличия предпочитаемых температур хорошо коррелируют с разницей температурных условий в естественных местообитаниях, показывая совпадение поведенческих реакций с настройкой физиологических механизмов температурных адаптации. [13]
Поясним эту закономерность несколькими примерами на материале, подробно изложенном в предыдущих главах. В системе теплообмена гомойотермных животных определенная группа приспособительных механизмов обеспечивает общий уровень адаптирован-ности организма к средним температурным условиям данного географического района и ( или) сезона года. Иногда эти приспособления объединяют термином температурные адаптации ( И. [14]
Прежде всего - и это самое основное - мы можем понять значение чрезвычайно высокого энергетического барьера для реакций, сопровождающихся образованием или разрывом кова-лентных связей. При обычных физиологических температурах скорости ферментативных реакций на 8 - 12 порядков выше, чем скорости аналогичных реакций без катализа. Таким образом, самая основная проблема температурной адаптации была разрешена в ходе эволюции путем выработки катализаторов - ферментов. [15]
Страницы: 1 2