Изменение - конформация - фермент
Cтраница 1
Изменение конформации фермента является существенной особенностью механизма индуцированного соответствия и может происходить также в деформационном механизме, поэтому возникает вопрос, каковы временнйе соотношения между такими конформационными изменениями и самим каталитическим процессом. С другой стороны, большинство структурных изменений белков, которые можно обнаружить физическими методами, в большинстве случаев протекает за время от нескольких секунд до нескольких часов, по-видимому, вследствие того, что скорость определяющей стадии ( которая сама по себе быстра) предшествуют неблагоприятные равновесные стадии. Такие относительно медленные конформационные изменения не могут происходить в каждом каталитическом обороте фермента, однако могут иметь значение для осуществления контроля ферментативной активности, как и в случае механизма индуцированного соответствия. [1]
Большая часть связывающей энергии расходуется на изменение конформации фермента. Таким образом, большая часть энергии связывания расходуется на обеспечение изменения конформации фермента от неактивной к активной форме, а оставшаяся часть проявляется в виде наблюдаемой энергии связывания. [2]
Трудно объяснить этот период индукции каким-либо другим механизмом, помимо изменения конформации фермента или химической реакцией с ферментом, которые протекают с константой скорости, меньшей, чем константа скорости, характеризующая число оборотов фермента. Существование таких медленных конформационных изменений приводит к серьезным сложностям при изучении ферментативной кинетики методами быстрого смешивания, поскольку состояние фермента в отсутствие субстрата может отличаться от состояния фермента в его присутствии и взаимное превращение этих форм фермента может быть сравнительно медленным по сравнению с максимальной скоростью превращения субстрата. [3]
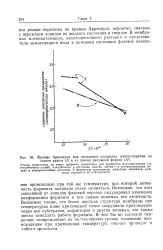 |
Кривые Аррениуса для окисления сукцината митохондриями из. [4] |
Возможно, что этот зависящий от липидов фазовый переход индуцировал изменение конформации фермента и тем самым понижал его активность. Возможно также, что более жесткая структура мембраны при температурах ниже критической точки затрудняла прохождение через нее субстратов, кофакторов и других молекул, что также могло замедлять работу фермента. В чем бы ни состояло первичное повреждение на молекулярном уровне, изменение, происходящее при критической температуре, обычно приводит к гибели организма. [5]
Зависимость автолиза от возраста связана не только с количеством имеющихся в клетке автолизинов, но и с их активностью, что обусловлено изменением конформации ферментов и физико-химических условий в стареющих клетках. [6]
Наиболее серьезная трудность в интерпретации изотопных эффектов ферментативных реакций заключается в том, что при замене воды на окись дейтерия существует определенная вероятность изменения конформации фермента вследствие изменений свойств водородных связей, гидрофобных взаимодействий или других факторов. Сами по себе эти эффекты малы, но они могут быть усилены в ферментативной реакции, в которой существует тонкое равновесие между конформациями различной степени активности. [7]
Большая часть связывающей энергии расходуется на изменение конформации фермента. Таким образом, большая часть энергии связывания расходуется на обеспечение изменения конформации фермента от неактивной к активной форме, а оставшаяся часть проявляется в виде наблюдаемой энергии связывания. [8]
Появление теории индуцированного соответствия было вызвано необходимостью объяснить некоторые факты, которые не удавалось согласовать с гипотезой ключа и замка. Известно, например, что в определенных ферментативных реакциях часть субстрата переносится на спирт, но не переносится на воду. Химические данные также указывают на изменение конформации фермента при связывании субстрата. [9]
Ферменты часто проявляют ингибирующее или активирующее влияние в присутствии физиологических концентраций метаболитов, которые являются предшественниками или продуктами метаболического пути, включающего данный фермент. Регулирование ферментативной активности по такому механизму обеспечивает поддержание концентраций метаболитов на физиологическом уровне. Такой контроль ферментативной активности может осуществляться изменениями конформации фермента, вызываемыми активаторами, ингибиторами или субстратами, и часто включает взаимодействия между субъединицами фермента. Особенно важными аспектами этой проблемы являются: 1) кооперативная природа таких взаимодействий и 2) контроль ферментативной активности посредством связывания молекулы с центром, отличающимся от активного центра. Изменения ферментативной активности, которые попадают в эту категорию, часто называют аллостерическими эффектами, однако использование этого термина, к сожалению, не ограничивается этим единственным смыслом. [10]
Субъединица ( 95 кДа) пронизывает мембрану насквозь и содержит на внутренней стороне участки для связывания АТФ и Na, а на внешней стороне - К и сердечных гликозидов. Субъединицы ( 40 кДа) содержат углеводные группы, расположенные на наружной стороне плазматической мембраны. Она способствует правильной ориентации фермента в липидном бислое. Перенос ионов происходит за счет изменения конформации фермента при его фрсфорили-ровании - дефосфорилировании за счет АТФ. [11]
В экспериментах по изучению равновесного связывания обнаружено присоединение одного или двух ионов фосфата на молекулу. Кроме того, Симпсон и Вейлли [76] получили кинетическое подтверждение подобной антикооперативности, обнаружив в соответствующих условиях активацию субстратом, выражающуюся в двухфазном характере зависимости активности от концентрации субстрата. Симпсон и Вейлли предполагают, что функциональная роль антикооперативности заключается в сохранении определенного уровня активности фермента в широком диапазоне концентрации субстрата. Их вывод о существовании в фосфатазе аллостерическнх взаимодействий отчасти подтверждается изменением конформации фермента при связывания ингибитора [70] [ ом. [12]
Страницы: 1