Структура - мембрана
Cтраница 1
 |
Схематическое изображение ультраструктуры мембран тилакоидов, собранных в граны, и мембран одиночных тилакоидов. [1] |
Структура мембраны как тилакоидов гран, так и тила-коидов стромы ( одиночных тилакоидов) была подробно изучена с помощью электронной микроскопии и иммунологических методов. Много информации получено также и о химическом составе этих мембран. [2]
 |
Схематическое изображение ультраструктуры мембран тилакоидов, собранных в граны, и мембран одиночных тилакоидов. [3] |
Структура мембраны асимметрична, ее липидный матрикс состоит из двух слоев, причем наружный слой толще внутреннего. В липидный матрикс погружено множество белковых частиц, которые видны как на наружной, так и на внутренней поверхности тилакоида. Они видны также по обе стороны от гидрофильной центральной зоны внутри мембраны на срезе препарата, приготовленного методом замораживания - скалывания. Эти частицы могут совершенно свободно перемещаться в жидком липидном матриксе. Их содержится до 6000 на 1 мкм2, и они занимают примерно 70 и 50 % площади тилакоидов гран и тилакоидов стромы соответственно. Фотохимическая активность, электронный транспорт и активность метаболических ферментов ассоциированы с частицами разных типов, и катализируемые ими реакции протекают в направлении, перпендикулярном к мембране. [4]
 |
Влияние длительности сушки при получении мембран из ароматического полиамида. [5] |
Структура мембраны и ее фильтрационные свойства определяются, главным образом, параметрами ее формования. Наиболее важными из них являются: продолжительность и температура сушки, температура и состав осадительной ванны, а также толщина слоя раствора. [6]
Исследование структуры мембран показало, что полученные образцы являются сплошными твердыми телами, не обладающими заметной капиллярной пористостью. При взаимодействии с водными растворами возникает вторичная нерегулярная пористость за счет микро - и ультрамикро-трещин, оказывающая влияние на электрохимическую активность таких мембран. [7]
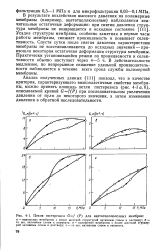 |
Петля гистерезиса Gf ( P для ацетатцеллюлозных мембран. [8] |
Усадка структуры мембраны, особенно заметная в первые часы работы мембраны, снижает проницаемость и повышает селективность. Спустя сутки после снятия давления характеристики мембраны не восстанавливаются до исходных значений - произошла некоторая остаточная деформация структуры мембраны. В действительности медленное, но непрерывное снижение удельной производительности наблюдается в течение всего срока службы полимерной мембраны. [9]
Нарушение структуры мембраны у мицелиальных грибов под влиянием ПАВ показано в работе ( 21) с помощью метода спиновой метки. [10]
Изучение структуры мембран митохондрий методами рант-гено-структурного анализа и электронной микроскопии позволило сделать заключение, что темные слои стенок мембраны ( рис. 3V) соответствуют - слоям белка, а более светлые - бимолекулярным слоям липоидов. Общая картина строения мембранных стенок митохондрий в настоящее время представляется такой, какой она - показана а рисунке 3V. Каждая мембрана состоит из двух слоев белковых молекул и заключенных между ними двух слоев липидов. На этих белковых и ли-пидных слоях адсорбированы ферменты, которые катализируют биохимические реакции в митохондриях. [11]
На структуру мембран значительное влияние оказывают следующие факторы: тип заряженной частицы, присутствие примесей в полимере, концентрация раствора, вид и длительность дополнительного облучения, частичный отжиг перед выщелачиванием, продолжительность химического травления. При частичном отжиге и применении низкоконцентрированного травильного раствора получаются мембраны с порами малого-диаметра. [12]
На структуру мембран значительное влияние оказывает скорость диффузионных процессов, переводящих систему в области, ограниченные бинодалью или спинодалью, а также скорость собственно разделения на фазы. [13]
Именно сжатием структуры мембраны и выдавливанием из нее части воды, происходящим под действием отрицательного давления, объясняется появление градиента концентрации, показанного на рис. П-49. При малой кривизне менисков ( давление пара под мембраной близко к давлению насыщения) градиент отрицательного давления, а следовательно, и концентрации мал, а при большой кривизне - градиенты велики. С течением времени градиент концентрации сильно меняется, однако это совершенно не сказывается на скорости испарения, которая практически остается постоянной. Его можно просто объяснить, основываясь на предложенном механизме проницания. Вода быстро проницает от одной поверхности к другой по крупным капиллярам, занимающим очень малую часть площади мембраны. Набухание же остальной ее части является медленным процессом и идет с двух сторон мембраны, так как они обе покрыты водой, по направлению к ее средним слоям. Если пренебречь капиллярным характером проницания и воспользоваться полученным распределением концентраций для вычисления коэффициента диффузии, то окажется, что коэффициент диффузии с ростом концентрации уменьшается. Некоторые авторы: этот факт объясняют глобулообразованием. Описанные выше опыты показывают, что его можно объяснить проще, если предположить, что течение через эти мембраны является капиллярным и диффузионный поток в этом случае незначителен. [14]
На податливость структуры мембран могут оказывать влияние некоторые добавки в грунтовку, которые для материала мембраны являются пластификаторами. Так, водные растворы триэтиламина и некоторых спиртов - пластификаторы для ацетатов целлюлозы. Поэтому повышение концентрации триэтиламина или спирта в грунтовке или в промывной воде выше определенной нормы будет ускорять пластическую деформацию мембран и может привести к резкому изменению ее эксплуатационных характеристик. Структурные превращения в мембране под действием давления, очевидно, происходят в макроструктуре и не захватывают тонкую надмолекулярную структуру ацетата целлюлозы. [15]
Страницы: 1 2 3 4