Структура - глобула
Cтраница 1
Структура глобулы окисленного цит с тождественна для лошади и рыб - бонито и тунца. [1]
Кроме того, структура малой глобулы гораздо менее универсальна, чем большой, и более зависима от структуры макромолекулы. [2]
Некоторые общие особенности структуры глобулы карбоан-гидразы заметны и на этих картах. На пространственной модели полипептидной цепи, построенной в соответствии с ними [23], обнаруживаются четыре фрагмента правой спирали, по два витка в каждом. Как отмечалось в разд. ДОВ и КД различных карбоангидраз также указывают на малую степень спирализации. [3]
Таким образом, вблизи точки перехода ( Л - - 1) структура глобулы и в общем случае напоминает рис. 1.12 - вокруг области локализации поля р ( х) флуктуируют длинные гауссовы петли. [4]
Из формулы (20.27) следует, что вблизи 9-температуры значение п0 действительно мало, так что для анализа структуры глобулы можно использовать ви-риальное разложение. Результат (20.27) показывает также, что равновесный объем макромолекулы в глобулярном состоянии вблизи 9-температуры устанавливается таким, чтобы притяжение ( слипание) звеньев при попарных контактах компенсировалось их отталкиванием при тройных столкновениях. [5]
Такой вывод нельзя считать окончательным - он опирается на ограниченный материал и не согласуется с общими представлениями о структуре глобулы. [6]
Ядро глобулы подобно кристаллу, а не жидкости, оно характеризуется нулевой конфигурационной энтропией. Самоорганизация структуры глобулы означает выбор одной наиболее выгодной микроструктуры. [7]
Допустим, что истинный активационный барьер для данной стадии ферментативного процесса слабо зависит от температуры, поскольку организованная глобула фермента ведет себя как кооперативная система. Иными словами, сама структура глобулы зависит от температуры. Допустим также, что при этом истинное значение S не зависит от температуры. [8]
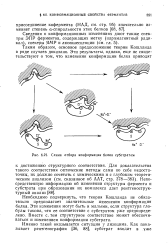 |
Схема отбора конформации белка субстратом. [9] |
Необходимо подчеркнуть, что теория Кошланда не обязательно предполагает значительные изменения конформации белка. Эти изменения могут быть и малыми, если структура глобулы такова, что ее соответствие с субстратом уже предопределено. [10]
В отличие от полярона, конформон не является истинной квазичастицей, способной перемещаться на большие расстояния. Его энергия быстро диссипирует из-за неоднородности и апериодичности структуры глобулы. Дальнейшая конформационная перестройка глобулы может происходить уже не в результате ЭКВ, но вследствие кооперативности макромолекулы. [11]
Из этого следует, что правильные замещения должны меньше сказываться на структуре и, следовательно, функции белка, чем неправильные. Правильные замещения, сохраняющие число и относительное расположение п - и н-остатков, должны меньше влиять на структуру глобулы, чем неправильные замещения, меняющие эти характеристики. [12]
Белковая цепь может иметь громадное число конформаций. Эта задача, по-видимому, обходится и природой, так как такой перебор потребовал бы очень большого времени, а самосборка белковой глобулы происходит за время порядка 1 с. Основная идея современных работ, посвященных предсказанию структуры глобулы, исходя из знания первичной структуры цепи, состоит в том, что нативная глобула есть конечный результат самосборки, не обязательно отвечающий абсолютному минимуму свободной энергии. [13]
Очевидно, что ферментативная активность должна существенно зависеть от рН среды. То же относится к большинству субстратов, модификаторов и коферментов. Изменение рН может изменить состояние ионизуемых групп в активном центре или соседствующих с ним; оно может также влиять на состояние неферментных компонент системы и на структуру глобулы локально или в целом. [14]
Между взаимодействием боковых радикалов и тенденцией полипептидной цепи образовывать спирали существует антагонизм и конкуренция. В тех местах, где спирали ломаются и изгибаются, регулярность их нарушена и образуются аморфные области макромолекулы. Под третичной структурой белка мы понимаем укладку спиральных и аморфных участков полипептидных цепей в компактном теле - глобуле. Силы, определяющие третичную структуру белка, это, с одной стороны, вандерваальсовы силы между боковыми радикалами, с другой стороны, дисульфидные и иные ковалентные связи, придающие жесткость структуре глобулы. [15]
Страницы: 1