Сульфатредуктор
Cтраница 2
Примерами крайних алкалофилов могут служить архебактерии Natronobacterium; уробактерии Sporosarciria urea; разлагающие мочевину с образованием аммиака, обитатели содовых солончаков Bacillus. В последнее время были открыты экстремально алкалофильные сульфатредукторы Desulfonatronovibrio и Desulfonatronum, гомоацетогенный галоанаэроб Natroniella, и ряд других организмов, принадлежащих к разным ветвям филогенетического дерева. Особо следует отметить многочисленность алкалофильных цианобактерий. [16]
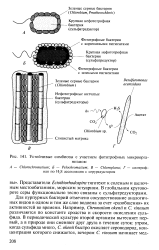 |
Устойчивые симбиозы с участием фототрофных микроорганизмов. [17] |
Представители Ectothiorhodospira тяготеют к соленым и щелочным местообитаниям, морским эстуариям. В глобальном круговороте серы функционально тесно связаны с сульфатредукторами. Для пурпурных бактерий отмечено сосуществование аналогичных видов в одном и том же слое водоема за счет разобщения их активностей во времени. [18]
От магнитотактических бактерий следует отличать анаэробные магнетит-образующие анаэробы, для которых образование магнетита служит катаболическим процессом. Часть из них, как упомянутый Geobactef, близки сульфатредукторам, но многие относятся к совсем другим группам. Среди анаэробных магнетит-образующих бактерий многие используют водород для восстановления ферригидрита и являются эффективными гидрогенотрофами. За-варзина, образование магнетита происходит у них вне клетки в процессе диагенеза из аморфного предшественника. [19]
Такие микроорганизмы подразделяют на свободноживущие и симбиотические. Свободноживущие азотфиксаторы относятся к хе-мотрофам ( это анаэробные клостридии и сульфатредукторы, факультативно анаэробные бациллы и представители сем. [20]
Характерным во взаимодействии сульфидогенного и метано-генного сообществ служит существование так называемого неконкурентного С ] - пути. Например, при разложении пектина образуется метанол, не используемый сульфатредукторами, но окисляемый метилотрофными метаногенами. Даже при избытке сульфата и подавлении других метаногенов, например, в гиперсоленых водоемах, метилотрофные метаногены продолжают действовать в неконкурентном пути метаногенеза и служить источником метана. Для простоты на схеме неконкурентный Сгпуть не показан. [21]
Полагаем, что необходимо определить влияние ингибиторов на ряд пластовых микроорганизмов, развивающихся в сообществе: аэробных и анаэробных. Такими организмами, в частности, являются наиболее распространенные в пластах псевдомонасы и сульфатредукторы. Культивирование посевов с добавками ингибитора рекомендуется проводить при температуре, близкой к температурным условиям пласта. Желательно получить предварительно накопительные культуры пластовых микроорганизмов. Кроме того, для испытаний действия ингибиторов должны быть подобраны музейные штаммы сульфатвосстанавливающих бактерий, относящихся к разным видам и родам. Разные представители этих бактерий отличаются различной устойчивостью к некоторым ингибиторам. [22]
Интересным макрообразованием является циано-бактериалъный мат. В его состав входят цианобактерии, пурпурные и зеленые бактерии, а также сульфатредукторы. [23]
При сульфатном дыхании конечным акцептором электронов служит сульфат. Физиологическую группу бактерий, восстанавливающих сульфат, называют сульфатредуцирующими, десульфа-тирующими или сульфидогенными. Сульфатредукторы широко распространены в таксономических группах, но все они, в отличие от нитратредукторов, строгие ( облигатные) анаэробы. Обычно обитают в морских донных осадках. [24]
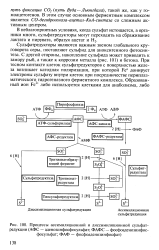 |
Процессы ассимиляционной и диссимиляционной сульфат. [25] |
Сульфатредукторы являются важным звеном глобального круговорота серы, поставляют сульфид для аноксигенного фотосинтеза. С другой стороны, накопление сульфида может приводить к замору рыб, а также к коррозии металла ( рис. 101) и бетона. При тесном контакте клеток сульфатредукторов с поверхностью железа возникает катодная поляризация, при которой Fe донирует электроны сульфату внутри клеток при посредничестве периплаз-матического гидрогеназного ферментного комплекса. [26]
Порядок использования Н2 и ацетата в анаэробном сообществе очень четкий: первым утилизируется Н2 и только с большой задержкой начинает использоваться ацетат. Это не столь отчетливо видно по значениям и тахдля чистых культур. Свободных осадках ацетат потребляется в основном сульфатредукторами в условиях избытка сульфата. В метантенках это не так ясно. В морских осадках Desulfobacter postgatei имеет большее сродство к ацетату, чем Methanosarcina barkeri. Но в метантенках метаносарцины работают только в переходный период, уступая место метанотриксам, которые имеют большее сродство к ацетату. При оценке конкуренции следует учитывать и начальное соотношение биомассы конкурирующих организмов. Особенно важно это для медленно растущих организмов. Находящиеся в избытке организмы могут получить преимущество даже при худших для данных условий кинетических характеристиках. [27]
Метаногены встречаются во всех анаэробных местах обитания, но в морских водоемах сульфатредукторы опережают их в борьбе за субстрат, так как эффективнее получают энергию. У сульфатредукторов ацетат окисляется через ЦТК и есть полная электронтранспортная цепь, заканчивающаяся сульфатом. В то же время многие морские метаногены способны использовать метиламины, которые не являются субстратами для сульфатредукторов. [28]
Конкуренция между использующими ЛЖК сульфатредуктора-ми и синтрофными организмами сложна. При высокой концентрации сульфата эти сульфатредукторы растут быстрее синтрофов и являются основными потребителями ЛЖК. При низкой концентрации сульфата возможно разложение ЛЖК синтрофами на ацетат и водород с конкуренцией за эти субстраты между метаноге-нами и сульфатредукторами. [29]
Это облигатно анаэробный организм, обладающий диссимиляторным путем восстановления сульфата с образованием H S. Он использует крайне ограниченное число субстратов, в первую очередь лактат, этанол, с неполным окислением их в ацетат. Сульфатредукторы способны к окислению Н2 и в этом случае осуществляют литоге-теротрофный рост. Комбинация субстратов Н2 ( или формиат), лактат, этанол наиболее обычна для большинства сульфатредук-торов. Анаболизм их основан на использовании ацетата с последующим карбоксилированием в трехуглеродное соединение. Ацетат как субстрат катаболизма использует Desulfobacter - - прямая палочка с заметно более медленным ростом, чем Desulfovibrio. [30]
Страницы: 1 2 3