Изменение - структура - белок
Cтраница 1
Изменение структуры белков в процессах денатурации обычно связано с К. [1]
В обоих случаях, кроме того, может иметь место кооперативное изменение структуры белка ( разд. Связывание с ионом металла может также ускорять процесс путем удерживания молекулы азота вблизи восстановительного центра. [2]
Для сложных молекул ( например, белков) изменение температуры может привести не только к изменению Больцмановского распределения молекул по уровням энергии, но и к изменениям структуры белка. S) и (4.8) могут оказаться неверными и использование уравнения Вант-Гоффа в термодинамике так же, как и Аррениуса и Эйринга в кинетике, могут привести к ошибочным результатам. [3]
Температурная зависимость спин-решеточной релаксации протонов белка в образце, содержащем 0 07 г D2O / r лизоцима, приведена на рис. 8.3. Возможно, здесь имеет место небольшое диполь-дипольное взаимодействие, обусловленное наличием дейтерия, и некоторое изменение структуры белка, вызванное заменой воды на дейтерий. Однако эти данные, по-видимому, надо рассматривать как удовлетворительную оценку величины Ню для гидратированного лизоцима. В ранее опубликованной работе [23], посвященной исследованию сухого лизоцима, получены сходные кривые, имеющие минимум вблизи 180 К. Как видно из этих данных, присутствие воды на белке приводит к существенному увеличению скорости релаксации. Не было сделано никакой попытки обнаружить релаксацию, описываемую с помощью двух экспонент; измерялась только медленная компонента релаксационной кривой. [4]
Белок может повысить скорость либо путем термодинамической стабилизации НО2 -, промежуточного продукта или конечного продукта реакции, либо обе стадии процесса или одна из них могут ускоряться за счет образования подходящих водородных связей, что, вероятно, связано также с кооперативными изменениями структуры белка. Но, поскольку неизвестны ни природа интермедиата, ни рН - зависимость каждой из стадий, ничего более конкретного об этом механизме сказать нельзя. [5]
Рассмотрим теперь, что дают однократные замещения нуклеотидов в кодонах. Наиболее опасны в смысле изменения структуры белка мутационные замещения в корне у. Кроме того, 7 замещений отвечают переходам б-о, 7 - переходам о-из п 2 - переходам 6-кбг. [6]
В статье II Денюэлем были уже рассмотрены основные реакции карбоксильных и амимных групп аминокислот, а также некоторые специфические реакции кислых, основных, ароматических, окси - и серусодержащих аминокислот. Химические реакции белков, очевидно, аналогичны реакциям этих функциональных трупп и лишь в незначительной степени изменяются с изменением структуры белка. По этой причине многие модельные реакции, проводимые на аминокислотах с целью определения специфичности белковых реагентов, в этой статье не упоминаются. Многообразные функциональные группы различных белков сгруппированы Тристрамом в таблицы ( статья III), и содержание их можно сравнивать с количеством введенного реагента. Здесь вряд ли стоит повторять сведения о химическом строении этих групп. [7]
Тем не менее появляющиеся примеры вызывают восхищение. В следующем десятилетии мы станем свидетелями новых усилий, направленных на изменение структуры ферментов, чтобы сделать их более полезными для промышленности, на изменение структур белков и пептидов для создания новых лекарственных препаратов. Ученые будут стремиться узнать больше о генетической регуляции и болезнях человека. [8]
Рассмотрим теперь, что дают однократные замещения нуклеотидов в кодонах. Наиболее опасны в смысле изменения структуры белка мутационные замещения в корне у. [9]
Цитоплазма обладает сложной прижизненной структурой, с которой связаны ее свойства и функции. Важнейшая из них - - избирательная проницаемость. Живая цитоплазма пропускает в вакуоль витальные красители, которыми клеточный сок и окрашивается. С гибелью или повреждением клетки в связи с изменением структуры белков красители задерживаются в самой цитоплазме. Цитоплазма и ядро приобретают соответствующую окраску. [10]
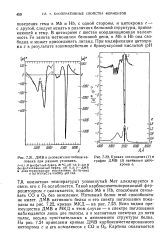 |
Кривая поглощения ( 1 и график ДМВ ( 2 нативного цитохрома с. [11] |
Мет алкилируется и связь его с Fe ослабляется. Такой карбоксиметилированный фер-роцитохром с оказывается, подобно Mb и НЬ, способным связывать СО и 02 без окисления. Нативный белок этой способности не имеет. ДМВ нативного белка и его спектр поглощения показаны на рис. 7.22, кривая МКД - на рис. 7.23. Ясно видны преимущества ДМВ и МКД в этом случае - в спектре поглощения наблюдаются лишь две полосы, а в магнитных спектрах до пяти полос, весьма чувствительных к изменениям структуры белка. [12]
Страницы: 1