Аденозиндифосфат
Cтраница 2
АТФ расходует свою богатую энергией связь, гидролизуясь до аденозиндифосфата ( АДФ) и фосфорной кислоты. Суммарно эту реакцию можно записать как АТФ-АДФ4-Ф - Однако при образовании фосфатов Сахаров эта энергия не расходуется на гидролиз, а используется для образования фосфатов за счет реакции переноса фосфорильной труппы на гидроксильные группы сахара, а не воды. [16]
Синтетический ацетилфосфат вступает в подобную реакцию только в присутствии аденозиндифосфата. В свете более поздних исследований представляется вероятным, что это новое вещество, так похожее на ацетилфосфат, является S-ацетиль-ным производным кофермента А. [17]
Это фосфорилирование идет при участии аденозинтрифосфата, превращающегося при этом в аденозиндифосфат. Вторая макроэргическая связь АТФ не может быть непосредственно использована, как источник энергии для химических синтезов или вообще для продуцирования энергии. Но и она становится доступной для использования вследствие деятельности фермента миокиназы. [18]
Главным фактором, контролирующим интенсивность дыхания живой ткани, является наличие аденозиндифосфата ( АДФ) и неорганического фосфата ( Фн), которые необходимы для образования аденозинтрифосфата ( АТФ) ( гл. Так как концентрация Ф редко бывает недостаточной, то интенсивность дыхания обычно контролируется наличием акцепторов фосфата, важнейшим из которых является АДФ. [19]
В этой главе введены следующие сокращения: AMP - адено-зинмонофосфат, ADP - аденозиндифосфат, АТР - аденозинтри-фосфат, DPN - дифосфопиридиннуклеотид, TPN - трифосфопи-ридиннуклеотид, DPNH - восстановленный дифосфопиридиннуклеотид, TPNH - восстановленный трифосфопиридиннуклеотид, иВРглюкоза - уридиндифосфатглюкоза, GMP - гуанозинмоно-фосфат, СМР - цитидинмонофосфат, UMP - уридинмонофос-фат, DHK. РНК - рибонуклеиновая кислота, IMP - инозинмонофосфат, ТМР - - тимидинмонофосфат, АВРглюкоза - аденозиндифосфатглюкоза, СВРманноза - гуанозиндифосфатманноза, СВРглюкоза - цити-динфосфатглюкоза, иВРглюкоза - уридиндифосфатглюкоза, иОРглюкуроновая кислота - уридиндифосфатуроновая кислота, иВРацетилглюкозамин - уридиндифосфат - Н - ацетилглюко-замин. [20]
Эфиры аденозина с фосфорной кислотой ( адениловая кислота), с пирофосфорной кислотой ( аденозиндифосфат, АДФ) и трифос-форной кислотой ( аденозинтрифосфат, АТФ) встречаются во всех живых клетках. [21]
Эфиры аденозина с фосфорной кислотой ( адениловая кислота), с пирофосфорной кислотой ( аденозиндифосфат, АДФ) и трйфосфорной кислотой ( аденозинтрифосфат, АТФ) встречаются во всех живых клетках. [22]
Найдено, что для реакций ( 3) и ( 4) требуется присутствие аденозиндифосфата и либо арсената, либо фосфата. Таким образом, синтез глутатиона из глутамилцистеина и глицина сходен в некоторых отношениях с синтезом глутамина ( стр. В то время как гидроксиламин может замещать глицин в реакции ( 2), что приводит к образованию гидроксамовой кислоты, другие аминокислоты и аммиак не активны в этой системе. [23]
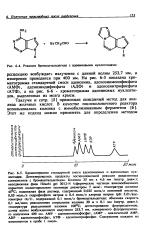
На рис. 6 - 5 показана хро-матограмма стандартной смеси аденозина, аденозинмонофосфата ( АМФ), аденозиндифосфата ( АДФ) и аденозинтрифосфата ( АТФ), а на рис. 6 - 6 - хроматограмма адениновых нуклеоти-дов, выделенных из мозга крысы. [25]
Энергия освобождается при экзотермической реакции гидроли - ва АТФ с отщеплением одной фосфатной группы и образованием аденозиндифосфата: энергетический эффект составляет 35 - 60 кДж / моль. [26]
Следует отметить, что наряду с АТР в живых организмах встречаются, но в значительно меньших количествах, аденозиндифосфат ( АДР), аденозинмо-нофосфат ( AMP), циклический адено-зинмонофосфат и диаденозинтетра-фосфорная кислота, играющие регуля-торную роль в обмене веществ. [27]
Энергия, накопленная в высокоэргических фосфатных связях 1 3-ди-фосфоглицериновой или фосфопировиноградной кислоты, без потерь переносится на молекулу аденозиндифосфата, который при этом превращается в аденозинтрифосфат. [28]
Энергия, накопленная в высокоэргических фосфатных связях 1 3-ди-фосфоглицериновой и фосфопировиноградной кислоты, без потерь переносится на молекулу аденозиндифосфата, который при этом превращается в аденозинтрифосфат. [29]
Все рассмотренные реакции протекают в присутствии разных ферментов с участием ряда коферментов, главнейшие из которых: аденозинтрифосфат, аденозиндифосфат, уридинглюкозодифосфат и др. При освещении в присутствии меченых фосфатов эти вещества быстро усваивают радиоактивный фосфор. Это подтверждает, что аденозинтрифосфат ( наряду с адено-зиндифосфатом и уридинглюкозодифосфатом) активно участвует в метаболизме фосфора при фотосинтезе. С другой стороны, увеличение содержания Рза в фосфороглицериновой кислоте при освещении указывает на связь ассимиляции фосфора с фотосинтезом, что раньше не удавалось достаточно убедительно доказать из-за экспериментальных затруднений. [30]
Страницы: 1 2 3 4