Тета-ритм
Cтраница 1
Тета-ритм может быть зафиксирован и во время активных фаз сна ( парадоксальный сон), которые сопровождаются частыми движениями глаз. [1]
Продолжительность вызванного тета-ритма характеризует его как реакцию на раздражение. [2]
Альфа - и тета-ритм условились называть быстрым сном, дельта-ритм - медленным. В течение ночи оба сна несколько раз сменяют друг друга ( полный цикл длится 1 5 часа), причем в последующих циклах доля быстрого сна возрастает и может достигать 30 и более минут. [3]
Далее нам удалось показать, что тета-ритм может установиться и без внешней причины как сопровождение истинного поискового и ориентировочного поведения, а также как следствие ожидания при отсутствии сигнала. [4]
Таким образом, мы можем предположить, что тета-ритм связан с активированием аммонова рога и специфическими, направленными на приобретение информации способами поведения. [5]
Авторы установили постепенное, от подкрепления к подкреплению, возрастание мощности тета-ритма дорсального отдела гиппокампа при доминировании одной из мотиваций и снижение ее после смены одной мотивации на другую. [6]
Если предсказание не подтверждается, то наступает сенсибильный период получения информации, сопоставления информации и внутренней корректуры модели, что объективируется потенциалами несоответствия или тета-ритмом. [7]
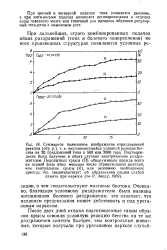
При третьей и четвертой подачах тона учащается дыхание, а при пяти-восьми подачах возникает десинхронизация в отделах коры головного мозга или типичный для процесса обучения регулярный тета-ритм в аммоновом роге. [9]
Юнг и Корнмюллер в 1938 г. открыли в точках мозга в области аммонова рога кролика постоянный ритм в 4 - 7 герц ( называемый также тета-ритмом), который затем был обнаружен у крыс, кошек и обезьян, когда животные настраивались на ориентировочное и учебное поведение. [10]
Хотя детальных сведений о результатах экспериментов авторы не приводят, можно предполагать, что низкочастотные ЭМП ( 2 - 10 Гц) вызывали преобладание колебаний биопотенциалов в диапазоне дельта - и тета-ритмов, а ЭМП 400 Гц - преобладание высокочастотных колебаний в бета-диапазоне. [11]
По мере развития техники или в каких-то экстраординарных ситуациях можно, конечно, использовать электроэнцефалограф, который будет определенным образом соединен с магнитофоном и, когда уловит появление в мозге ритма в пределах 4 - 7 колебаний в секунду ( так называемый тета-ритм, при котором только и возможно успешное обучение), то включит магнитофон. Вероятно, это не за горами, однако сегодня лучше не уповать на чудеса, а по старинке, пусть в 2 - 3 раза медленнее, учиться в состоянии бодрствования. [12]
Если ее параметры изменяются, то возникают импульсы несоответствия, вызывающие ориентировочное поведение. Суммарные потенциалы тета-ритма, по всей вероятности, не служат причиной ориентировочных движений, а представляют собой сопутствующее нервным процессам явление. Целенаправленное разведывание дифференцирует нейрональ-ную память и благодаря процессу привыкания приводит к угасанию ориентировочной реакции и исчезновению соответствующих потенциалов. [13]
К сожалению, критерии для оценки эмоционального напряжения в периоде ожидания очень не конкретны. Целый ряд физиологических показателей ( сдвиг волн ЭЭГ в сторону более высоких частот, увеличение индекса тета-ритма, увеличение амплитуды вызванных потенциалов, изменение некоторых вегетативных показателей) не всегда может доказать степень эмоционального напряжения человека в период ожидания. Эта неопределенность обусловлена тем, что в периоде ожидания возникает дезинтеграция приспособительных регуляторных механизмов в самых разнообразных системах: психической, вегетативно-гормональной, функционально-моторной. Кроме того, большую роль играют такие базисные факторы, как интеллектуальное развитие, различные генетические детерминанты, возраст и характер предыдущего жизненного опыта. От реакции всех этих систем в период ожидания зависит надежность в выполнении оператором его функциональных обязанностей. Учитывая такое многоплановое отраженна процесса ожидания в различных психофизиологических коррелятах, мы в своей деятельности изучали этот процесс с помощью различных тестов: ЭЭГ, РЭГ, ЭКГ, КГР, определение катехолами-нов в моче и 17-кетостероидов. Именно суммарная оценка динамических сдвигов этих показателей позволяет не только с качественной, но и количественной стороны оценить напряженность процесса ожидания и дать неспецифическую характеристику общего тонуса нервной системы. [14]
Различают локально-модулирующие и генерализованно-модулирующие нейроны. Важный класс модулирующих нейронов составляют ритмические модуляторы: нейроны неспецифического таламуса, ответственные за генерацию аль-фаподобных колебаний, и нейроны септума, ответственные за генерацию тета-ритма. [15]
Страницы: 1 2