Точка - расщепление
Cтраница 2
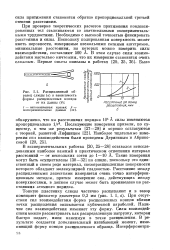 |
Расщепленный разец слюды ( а и зависимость формы расщепленных концов от их длины ( б. [16] |
Тонкую пластинку слюды частично расщепляют и в зазор помещают фиксатор диаметром 0 3 мм ( рис. I. При отсутствии сил взаимодействия форма расщепленных концов вблизи точки расщепления описывается кубической параболой. Наличие сил взаимодействия изменяет эту форму. Силы взаимодействия можно рассматривать как распределенную нагрузку, которая быстро падает, имея максимум в точке расщепления. В результате создается дополнительный изгибающий момент, изменяющий форму концов расщепленного образца. [17]
Имеющиеся эксперименты указывают на то, что в ряде случаев детонационная волна при ослаблении не вырождается в волну Чепмена-Жуге, а на некотором расстоянии от тела расщепляется на адиабатическую ударную волну и фронт медленного горения. В рамках теории детонационного фронта нулевой толщины положение точки расщепления не может быть определено [1] и для его определения необходимо рассматривать структуру зоны горения в детонационной волне. [18]
Как показывают приведенные выше результаты, при достаточно большом тепловыделении сильная детонационная волна перед телом очень быстро стремится к детонации Чепмена-Жуге. Поэтому, если в таких случаях происходит расщепление волны, то точка расщепления будет находиться на небольшом расстоянии от тела. Найденные в работе [4] два асимптотических режима обтекания поджигающего источника соответствуют двум возможным случаям: в первом - детонационная волна не расщепляется, постепенно ослабевает при удалении от тела и превращается в волну Чепмена-Жуге, во втором - она расщепляется, и на больших расстояниях от тела интенсивность ударной волны определяется распространяющимся по смеси фронтом медленного горения. [19]
Эти ацетальные связи устойчивы к щелочи, даже при жестких условиях, но легко гидролизуются кислотами. Каждый раз, когда кислоты расщепляют глюкозидную ( ацетальную) связь, они образуют новую редуцирующую и новую гидроксильную группы в четвертом положении глюкозной единицы по другую сторону от точки расщепления. По мере того как средняя длина цепи у гидроцеллюлоз становится короче, число концевых групп увеличивается, и гидроцеллюлозы почти во всех отношениях проявляют присущие им свойства. Опыт с редуцирующими сахарами наводит на мысль, что эти альдегидные группы существуют как циклические полуацетали ( структура 5, RH), а не в обычной альдегидной форме. Эту точку зрения подтверждает наличие группировки СНО и их несколько аномальное поведение с реагентом Шиффа и другими. [20]
В схеме Годунова, в которой по параметрам на слое t из решения задачи о распаде произвольного разрыва находятся нормальные компоненты скорости центров всех элементов волны, построение контура волны можно вести аналогичным образом. При этом роль скорости звука играет своя для каждого элемента нормальная скорость D, а набегающий поток может быть и не равномерным. Для случая с точкой расщепления d соответствующая схема дана на рис. 2, в. Здесь kd - линия стационарного косого скачка, а тонкие прямые - направляющие разностной сетки. Поскольку в действительности для определения координат узлов строить штриховую и штрихпунктирную кривые не требуется, то алгоритм счета получается весьма простым. [21]
Действительно, в более тонких пленках ( см рис. 12) и в случае больших парциальных давлений кислорода ( см. рис. 13, - а) скорости и степень автокатализа выше. Совпадающие участки кривых 0о2 - - т ( см. рис. 12) для пленок, различающихся по толщине, соответствуют состоянию реакционной среды, когда скорость потребления кислорода даже в самых глубоких слоях более толстой пленки еще не достигла о2Пред и. В этом режиме стационарная концентрация кислорода во всех слоях пленки практически равна его растворимости в субстрате. Точки расщепления совпадающих участков кривых соответствуют переходу реакции в нижних слоях пленки толщиной 35 мкм в диффузионный режим. Положение точек расщепления и максимумов на кривых vо2 - т зависит от строения ОЭА и условий пленкообразования. [22]
Тонкую пластинку слюды частично расщепляют и в зазор помещают фиксатор диаметром 0 3 мм ( рис. I. При отсутствии сил взаимодействия форма расщепленных концов вблизи точки расщепления описывается кубической параболой. Наличие сил взаимодействия изменяет эту форму. Силы взаимодействия можно рассматривать как распределенную нагрузку, которая быстро падает, имея максимум в точке расщепления. В результате создается дополнительный изгибающий момент, изменяющий форму концов расщепленного образца. [23]
Локализация сайтов модификации в нуклеиновых кислотах обычно проводится методом, сходным с тем, который применяется при секвенированип ДНК методом Максама-Гилберта. Нуклеиновая кислота ( мишень) метится 32Р по одному из концов. После аффинной модификации реакциопноепоеобпым производным антисмыслового олигонуклеотида мишень обрабатывают так, чтобы разрушить межнуклеотидную связь в модифицированном сайте. На радиоавтографе, полученном после электрофоретического разделения, появляются полосы, соответствующие фрагментам мишени, расположенным между 32Р - мечеиным концом и точкой расщепления. Положение полосы соответствует длине фрагмента и, следовательно, указывает на расстояние между точкой расщепления и меченым концом и тем самым указывает на положение модифицированного остатка. В качестве примера на рис. 95 представлены, результаты аффинного алкнлированпя одноцепочечиой ДНК, состоящей из 303 нуклеотидов. [24]
Действительно, в более тонких пленках ( см рис. 12) и в случае больших парциальных давлений кислорода ( см. рис. 13, - а) скорости и степень автокатализа выше. Совпадающие участки кривых 0о2 - - т ( см. рис. 12) для пленок, различающихся по толщине, соответствуют состоянию реакционной среды, когда скорость потребления кислорода даже в самых глубоких слоях более толстой пленки еще не достигла о2Пред и. В этом режиме стационарная концентрация кислорода во всех слоях пленки практически равна его растворимости в субстрате. Точки расщепления совпадающих участков кривых соответствуют переходу реакции в нижних слоях пленки толщиной 35 мкм в диффузионный режим. Положение точек расщепления и максимумов на кривых vо2 - т зависит от строения ОЭА и условий пленкообразования. [25]
Локализация сайтов модификации в нуклеиновых кислотах обычно проводится методом, сходным с тем, который применяется при секвенированип ДНК методом Максама-Гилберта. Нуклеиновая кислота ( мишень) метится 32Р по одному из концов. После аффинной модификации реакциопноепоеобпым производным антисмыслового олигонуклеотида мишень обрабатывают так, чтобы разрушить межнуклеотидную связь в модифицированном сайте. На радиоавтографе, полученном после электрофоретического разделения, появляются полосы, соответствующие фрагментам мишени, расположенным между 32Р - мечеиным концом и точкой расщепления. Положение полосы соответствует длине фрагмента и, следовательно, указывает на расстояние между точкой расщепления и меченым концом и тем самым указывает на положение модифицированного остатка. В качестве примера на рис. 95 представлены, результаты аффинного алкнлированпя одноцепочечиой ДНК, состоящей из 303 нуклеотидов. [26]
Очевидно, что N-концевые группы всех Т - пептидов отличаются от N-концевых групп С-пептидов, поскольку использованные для расщепления ферменты действуют по разным точкам. Исключение составляют пептиды, полученные из N-конца исходной цепи, они должны иметь одинаковое начало. Из рассмотрения приведенных в табл. 7.4 структур видно, что таковыми являются пептиды Т-10 и С-5. Следовательно, пептид серии Т, примыкающий с С-конца к Т-10, должен начинаться с фенилаланина. Таковым в приведенной серии является только пептид Т-4, т.е. последовательность трипсиновых фрагментов с N - сонца молекулы Т-10, Т-4. Этот двойной Т - пептид содержит весь пептид С-5 и сверх того фрагмент ER. Следовательно, к С-5 должен примыкать пептид С-7, начинающийся с этих двух аминокислотных остатков. Следующая за аргинином основная часть пептида С-7 является N-концевой частью пептида Т-14, который примыкает в исходной структуре к Т-4. Восстановленная таким путем N-концевая последовательность рибонуклеазы приобретает вид Т-10, Т-4, Т-14. Последний содержит остаток тирозина ( Y), т.е. точку расщепления химотрипсином. [27]
Страницы: 1 2