Транспозон
Cтраница 3
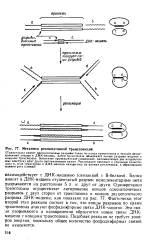 |
Механизм репликативной транспозиции. [31] |
Транспозаза вносит одноцепочечные разрывы точно по концу транспозона и косой двуце-почечный разрыв в ДНК-мишень. Затем транспозаза объединяет концы разрыва мишени с концами транспозона. Возникает промежуточное соединение, напоминающее две направленные навстречу друг другу реплнкативные вилки. Репликация приводит к удвоению транспо-лоиа и, если траиспозон и ДНК-мишень находились на разных репликонах. [32]
ДНК, в которой в результате вырезания транспозона остался разрыв, однако существуют и другие возможности. [33]
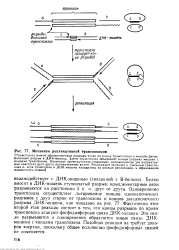 |
Механизм реплнкативнон транспозиции. [34] |
Транспозаза вносит одиоцепочечные разрывы точно по концу транспозона и косой двуце - Почечный разрыв в ДНК-мишень. Затем транспозаза объединяет концы разрыва мишени с концами транспозона. Возникает промежуточное соединение, напоминающее две направленные навстречу друг другу репликативные вилки. [35]
Однако стратегия выживания плазмид, как и в случае транспозонов, основывается не только на том, чтобы приносить пользу клеткам-хозяевам. Прежде всего сохранению плазмид в популяциях бактерий способствует высокая стабильность их наследования. Она достигается точным контролем репликации и секрегации плазмид по дочерним клеткам ( см. гл. II), но многие плазмиды имеют еще дополнительные приспособления для повышения стабильности наследования. Другая плазмида, R1 ( кодирующая устойчивость к нескольким антибиотикам), достигает той же цели другим путем. Один из генов этой плазмиды кодирует токсичное для клетки вещество, а другой ген - короткоживущий ингибитор этого вещества, так что наличие плазмиды для клетки не вредно. [36]
Однако стратегия выживания плазмид, как и в случае транспозонов, основывается не только на том, чтобы приносить пользу клеткам-хозяевам - Прежде всего сохранению плазмид в популяциях бактерий способствует высокая стабильность их наследования. Она достигается точным контролем репликации и секрегации плазмид по дочерним клеткам ( см. гл. II), но многие плазмиды имеют еще дополнительные приспособления для повышения стабильности наследования. Другая плазмида, R1 ( кодирующая устойчивость к нескольким антибиотикам), достигает той же цели другим путем. Один из генов этой плазмиды кодирует токсичное для клетки вещество, а другой ген - короткоживущий ингибитор этого вещества, так что наличие плазмиды для клетки не вредно. [37]
Ясно, что описанный механизм транспозиции годится не для всех транспозонов. Например, составные транспозоны, состоящие из какого-либо гена, окруженного IS-элементами в прямой взаимной ориентации, по структуре неотличимы от коинтеграта ( ср. Далее, многие транспозоны образуют коинтеграты, отличающиеся исключительной стабильностью. [38]
На основании полного восстановления активности му-тантных генов при вырезании IS-элементов и транспозонов предполагают, что вырезание происходит абсолютно точно и что дуплици-рованный участок ДНК-мишени снова оказывается представленным одной копией. В принципе, такой результат может быть достигнут за счет гомологичной рекомбинации между окружающими транспо-зон дуплицированными последовательностями или за счет ошибки репликации, вызванной наличием на концах транспозонов инвертированных повторов. [39]
Фактически этот второй этап реакции состоит в том, что концы разрывов по краям транспозона атакуют фосфодиэфирные связи ДНК-мишени. Эти связи разрываются и одновременно образуются новые связи ДНК-мишени с концами транспозона. Подобная реакция не требует доноров энергии, поскольку общее количество фосфодиэфирных связей не изменяется. [40]
Фактически этот второй этап реакции состоит в том, что концы разрывов по краям транспозона атакуют фосфодиэфирные связи ДНК-мишени. Эти свя -: зи разрываются и одновременно образуются новые связи ДНК-мишени с концами транспозона. Подобная реакция не требует доноров энергии, поскольку общее количество фосфодиэфирных связей не изменяется. [41]

Он обладает типичными свойствами фага, и в то же время его можно рассматривать как гигантский транспозон. [43]
По аналогичному механизму перемещается и транспозон ТпЮ, но в этом случае показано, что вырезавшийся транспозон образует кольцевую молекулу, видимо, способную затем встраиваться на новое место. Образование кольцевой ДНК еще раз указывает на то, что транспозазы одновременно взаимодействуют с двумя концами транспозона или IS-элемента. [44]
 |
Две основные реакции, осуществляемые IS - и Тп-элементами. [45] |
Страницы: 1 2 3 4