Триозофосфат
Cтраница 3
Таким образом, из трех молекул рибулозо-5 - фосфата образуются две молекулы гексозо-6 - фосфата и молекула триозофосфата; две молекулы триозофосфата дают еще одну молекулу глюкозо-6 - фосфата. Это соответствует стехиометрическому уравнению ( Д) ( см. стр. Суммирование этого уравнения с уравнением ( Е) для первых стадий процесса приводит к общему уравнению ( Г) ( см. стр. [31]
Карбоксильный же углерод уксусной кислоты распределяется поровну между СО2 ( образованным во время реакции катализируемой фос-фоенолпируваткарбоксикиназой) и триозофосфатом. [32]
Хлоропласт считают центром фотосинтеза, способным катализировать отдельно световую и темновую реакции и тем самым воздействовать на восстановление углекислоты до триозофосфата и фосфатов Сахаров. В то время как скорость световой реакции ( синтез АТФ и восстановление НАДФ), катализируемой изолированными хлоропластами, более чем достаточна для фотосинтеза in vivo ( см. выше), суммарная скорость ассимиляции углекислоты изолированными хлоропластами низка. Например, изолированные хлоропласта катализируют ассимиляцию со скоростью приблизительно 4 мкмоль углекислоты на 1 мг хлорофилла в 1 час, а в целой ткани этот процесс протекает со скоростью 200 мкмоль углекислоты на 1 мг хлорофилла в 1 час. Причина этого различия неясна. [33]
Таким образом, из трех молекул рибулозо-5 - фосфата образуются две молекулы гексозо-6 - фосфата и молекула триозофосфата; две молекулы триозофосфата дают еще одну молекулу глюкозо-6 - фосфата. Это соответствует стехиометрическому уравнению ( Д) ( см. стр. Суммирование этого уравнения с уравнением ( Е) для первых стадий процесса приводит к общему уравнению ( Г) ( см. стр. [34]
Образовавшийся фруктозо-6 - фосфат участвует в реакциях, характерных для пентозного цикла - При взаимодействии с ( 3 - й) молекулой триозофосфата и при участии транскетола-зы фруктозо-6 - фосфат превращается в ксилулозо-5 - фосфат и эритрозо-4 - фосфат. [35]
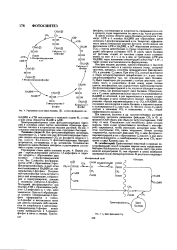 |
Упрощенна схема цикла Калвина. - остаток фосфорной к-та. [36] |
Эта Сз-кислота фосфорили-руется АТФ с образованием 3-фос-фоглицероилфосфата, к-рый затем СН2 восстанавливается НАДФН до гли-церальдегид-3 - фосфата. Полученный триозофосфат затем вступает в ряд р-ций изомеризации, конденсации и перегруппировок, дающих 3 молекулы рибулозо-5 - фосфата. Одна из 6 образующихся молекул глицеральдегид-3 - фос-фата превращается в глюко-зо-6 - фосфат и используется затем для синтеза крахмала либо выделяется из хлоропласта в цитоплазму. Глицеральдегид-3 - фосфат может также превращаться в 3-глицеро-фосфат и затем в липиды. [37]
 |
Схема фотосинтетического цикла. [38] |
Карбоксилирование D-рибулозо - 1 5-бисфосфата с последующим расщеплением промежуточного соединения на две молекулы 3-фосфоглицериновой кислоты является ключевой реакцией, поэтому такой фотосинтетический цикл называют С3 - путем, или С3 - типом фотосинтеза. Из этих триозофосфатов образуется D-фруктозо - 6-фосфат, часть которого выводится из цикла в качестве основного продукта фотосинтеза, а остаток вместе с неизрасходованными триозофосфатами идет на регенерацию D-рибулозо - 1 5-бисфосфата. Некоторые из промежуточных соединений также выводятся из цикла и наряду с D-фруктозо-б - фосфатом используются для синтеза различных углеводов и их производных. [39]
 |
Включение СО2 в различные промежуточные продукты метаболизма. [40] |
Сравнение всех трех типов автотрофной фиксации СО2 позволяет заключить, что анаэробные процессы более экономны, чем аэробные. Синтез 1 моля триозофосфата из 3 молей СО2 по анаэробному ацетил - СоА - пути требует затраты всего лишь 3 молей АТР, в восстановительном цикле трикарбоновых кис-лот-затраты 5 молей АТР, а в рибулозобисфосфатном цикле-9 молей АТР. [41]
Этот процесс сопровождается освобождением примерно V3 всей доступной свободной энергии. При разрушении углеводов образуются триозофосфаты и ( или) пируват; при разрушении жиров - ацетил - КоА ( см. гл. XIII), пропионил - КоА и глицерин; при разрушении белков - аце-тил - КоА, оксалоацетат, а-кетоглутарат, фумарат и сукцинат. Те же соединения образуются и тогда, когда организм - как это бывает у бактерий - использует не совсем обычные источники углерода, например некоторые ароматические соединения ( бензойная или миндальная кислота), алифатические ( у-аминомасляная или итаконовая кислота) и гетероциклические ( пурины или мочевая кислота) ( фиг. [42]
Центральные пути обмена веществ включают относительно небольшое число превращений. В основном это следующие реакции: триозофосфат пируват; пируват ацетил - КоА; оксалоацетат аспартат; а-кето ( оксо) - глутарат глутамат, а также циклическая система реакций, в ходе которых ацетил - КоА ( С2 на фиг. Последний механизм, известный под названием цикла лимонной кислоты ( ЦЛК), играет столь важную роль в организме, что описанию его посвящена особая глава ( гл. [43]
Мы уже видели, что у С6 - сахаров метятся два центральных атома углерода ( в положениях 3 и 4), а у триоз метится их нефосфорили-рованный концевой атом углерода. Таким образом, только путем конденсации триозофосфата с углеродными атомами 1 4 или 3 6 гексоз может образоваться седогептулоза, имеющая экспериментально найденное распределение метки. [44]
Фруктозобисфосфат дефосфорилируется под действием фруктозобис-фосфатазы с образованием фруктозо-6 - фосфата. Затем из одной молекулы фруктозо-6 - фосфата и трех молекул триозофосфата образуются три молекулы рибозо-5 - фосфата. Некоторые из ферментов, катализирующих эти превращения, участвуют и в окислительном пентозофос-фатном цикле. Транскетолаза катализирует перенос гликольной группы от кетозомон о фосфата на альдозофосфат. Гликолевый альдегид при этом временно связывается с тиаминдифосфатом ( тиаминпирофос-фатом), играющим роль кофермента: получается активный гликолевый альдегид. [45]
Страницы: 1 2 3 4