Терминирующий триплет
Cтраница 1
Терминирующий триплет в рамке считывания может появиться в кодирующей части мРНК в результате мутации. [1]
 |
Трансляционные стратегии вирусных ( РНК-геномов. [2] |
РНК; волнистые - полипептидные цепи: треугольники - ииицнаторные триплеты; квадраты - терминирующие триплеты ( черные - реализуемые в данной молекуле РНК. [3]

О терминации полипептида сигнализирует один из трех терминирующих кодонов мРНК, расположенный непосредственно за кодовом последней аминокислоты. Терминирующие триплеты UAA, UAG и UGA не кодируют никакую аминокислоту. Первоначально они были обнаружены при исследовании изменения одного-единственного нуклеотида в некоторых кодонах, соответствующих определенным аминокислотам. [5]
Так, инициирующий кодон AUG полиовирусной РНК расположен на расстоянии 743 нуклеотида от 5 -конца, и ему предшествуют восемь AUG триплетов во всех трех фазах считывания. Эти предшествующие AUG триплеты, однако, находятся не в подходящем структурном окружении ( см. выше), а кроме того, вслед за большинством из них на небольших расстояниях имеются терминирующие триплеты, прерывающие трансляцию, если бы она была на них инициирована. [6]
Надо сказать, что вне фазы ( рамки) считывания триплеты UAA, UAG и UGA в пределах кодирующей последовательности мРНК встречаются существенно чаще, чем в фазе считывания, где имеется, как правило, всего один терминирующий кодон на всю кодирующую последовательность. Поэтому обычно случайный сдвиг рамки в процессе элонгации не может привести к синтезу очень длинного неправильного полипептида и чаще всего приводит к скорой терминации этой неправильной трансляции. В некодирующих участках мРНК, включая межцистронные участки поли-цистронных РНК, частота терминирующих триплетов обычно также высока. [7]
Транскрипция экзонов и нитронов сложного эукариотического гена осуществляется в порядке их расположения друг за другом ( коллинеарная. Порядок вырезания районов нитронов необязательно соответствует последовательности их расположения в гене. Сплайсинг должен быть очень точным, в противном случае ошибка на один нуклеотид может вызвать нарушение рамки трансляции, что приводит, например, к образованию терминирующих триплетов и остановке синтеза полипептида. Однако границы экзонов и нитронов часто не удается определить с точностью до нуклеотнда, поскольку в районе сайтов сплайсинга нуклеотнды ( например, G, см. рис. 102) могут быть повторены. [8]
Транскрипция экзонов и интронов сложного эукариотического гена осуществляется в порядке их расположения друг за другом ( коллинеарнаятранскрипция, после чего вырезаются районы интронов, а экзоны сшиваются в результате сплайсинга. Порядок вырезания районов интронов необязательно соответствует последовательности их расположения в гене. Сплайсинг должен быть очень точным, в противном случае ошибка на один нуклеотид может вызвать нарушение рамкн трансляции, что приводит, например, к образованию терминирующих триплетов и остановке синтеза полипептида. Однако границы экзонов и интронов часто не удается определить с точностью до нуклеотида, поскольку в районе сайтов сплайсинга нуклеотнды ( например, G, см. рис. 102) могут быть повторены. [9]
Уже говорилось, что факторы терминации узнают терминирующий кодон, установленный именно в А-участке рибосомы. Прежде всего, кодонзависимое связывание RF возможно только после транслокации пептидил-т РНК из А-участка в Р - участок. В экспериментах in vitro можно прямо связать триплет AUG и инициатор-ную F-Met - tRNA с Р - участком вакантной рибосомы, а затем добавить RF и триплет UAA ( или UAG, или UGA); последние связываются и индуцируют гидролиз F-Met - tRNA в Р - участке с освобождением свободного формилметионина. Довольно очевидно, что в указанном случае терминирующий триплет и RF попадают в вакантный А-участок. Тот факт, что супрессорная аминоацил-т РНК с EF-TU и ГТФ, несущая антикодон, комплементарный терминирующему кодону, конкурирует с RF в процессе трансляции, тоже говорит об А-участке как месте связывания RF. Такие ингибиторы А-участка рибосомы, как тетрациклин и стрептомицин, ингибируют кодонзависимое связывание факторов терминации с рибосомой. EF-G, стабильно связанный с рибосомой ( например, в присутствии фусидовой кислоты), мешает как связыванию комплекса Aa - tRNA EF-TU GTP, так и RF с рибосомой. Белок прокариотической рибосомы L7 / L12, образующий боковой стержень на 50S субчастице, способствует как связыванию факторов элонгации, так и RF. Данные по ингибированию кодонзависимого связывания RF антителами к индивидуальным рибосомным белкам подтверждают, что место связывания RF сильно перекрывается одновременно с А-участком и участком связывания факторов элонгации. [10]
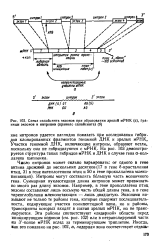 |
Схема сплайсинга экзонов при образовании зрелой мРНК ( а, границы экзоиов и нитронов ( правило сплайсинга ( б. [11] |
Интроны могут составлять большую часть мозаичного гена. Суммарная нуклеотидная длина нитронов может превышать во много раз длину экзонов. Например, в гене проколлагена птиц экзоны составляют лишь восьмую часть общей длины гена, а в гене тиреоглобулина млекопитающих - лишь двадцатую. Экзонами называют не только те районы гена, которые содержат последовательности, кодирующие участки полипептидной цепи, но и районы, транскрипты которых входят в состав зрелых мРНК, но не транслируются. Эти районы соответствуют лидерной области перед инициирующим кодовом ( см. рис. 102) или в нетранслируемой части на 3 -конце мРНК вслед за терминирующим триплетом. [12]
 |
Схема сплайсинга экзонов при образовании зрелой мРНК ( а, границы экзоиов и нитронов ( правило сплайсинга ( б. [13] |
Интроны могут составлять большую часть мозаичного гена. Суммарная нуклеотидная длина интронов может превышать во много раз длину экзонов. Например, в гене проколлагена птиц экзоны составляют лишь восьмую часть общей длины гена, а в гене тиреоглобулина млекопитающих - лишь двадцатую. Экзонами называют не только те районы гена, которые содержат последовательности, кодирующие участки полипептидной цепи, но и районы, транскрипты которых входят в состав зрелых мРНК, но не транслируются. Эти районы соответствуют лидерной области перед инициирующим кодоном ( см. рис. 102) или в нетранслируемой части на 3 -конце мРНК вслед за терминирующим триплетом. [14]
 |
Схема сплайсинга экзонов при образовании зрелой мРНК ( а, границы экзоиов и нитронов ( правило сплайсинга ( б. [15] |
Страницы: 1 2