Частичное убежище
Cтраница 1
Частичные убежища способствуют стабилизации динамики численности популяции хищника и жертвы. [1]
Такие частичные убежища также способствуют поддержанию устойчивости динамики взаимодействующих популяций. [2]
Существование частичных убежищ при взаимодействии хищник-жертва ведет к стабилизации динамики численности популяций. Даже при высокой плотности популяции хищника некоторое число жертв остается нетронутым, что защищает популяцию жертвы от наиболее сильных воздействий хищника. Это приводит к тому, что хищник всегда имеет в своем распоряжении популяцию жертвы, которой он питается. Обратите внимание на то, что такой стабилизирующий эффект является всего лишь следствием предпочтения, которое хищник оказывает наиболее выгодным кормовым пятнам; хищники ведут себя таким образом не для того, чтобы стабилизировать взаимодействия хищник-жертва. [3]
Наличие частичных убежищ ведет к тому, что изоклины жертвы становятся вертикальными при ее низкой плотности. А это приводит к низкой стабильной численности жертвы, когда хищник эффективен в поисках жертвы. Неоднородность среды, следовательно, оказывает стабилизирующий эффект, как это было показано ранее. [4]
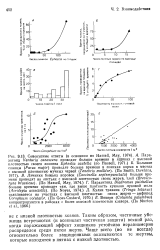
Таким образом, частичные убежища встречаются ( и возникает частичная защита) всякий раз, когда поражающий эффект хищников устойчиво неравномерно распределен среди пятен жертв. Чаще всего ( но не всегда) относительно более защищенными оказываются те жертвы, которые находятся в пятнах с низкой плотностью. [6]
Как уже было показано, функциональный ответ типа 3 дает такой же эффект, как и частичное убежище: низкая скорость выедания при низкой плотности жертвы. Однако в однородной среде существует противоречие между поведением хищника в соответствии с функциональным ответом типа 3 ( игнорирование жертв при низкой плотности) и высокой эффективностью добывания пищи при низкой плотности жертв. В этой ситуации будет применима кривая II на рис. 10.8 и стабилизирующее влияние ответа типа 3 ( разд. [7]

Агрегирующий ответ обычно приводит к параллельному возникновению неблагоприятных эффектов, но не всегда. Частичные убежища возникают в результате пятнистого распределения неблагоприятных эффектов. [9]
Растительноядные организмы часто образуют скопления, не являющиеся следствием агрегирующего ответа. Скопления растительноядных создают частичные убежища для растений. [10]
Личинки Cactoblastis агрегированы вдвойне: яйца откладываются кучками по 80 штук в каждой и сами кучки размещены по растениям агрегированно. Таким образом, это взаимодействие стабилизировано не только при низкой плотности тем, что кактусы в результате агрегированности личинок получают частичное убежище, но оно также стабилизировано благодаря высокой смертности личинок на чрезмерно зараженных растениях при высоком уровне численности личинок. [11]
Мы можем это сделать в рамках модели Лотки-Вольтер - ры с учетом того, что различные типы неоднородности обусловливают непропорционально пониженную скорость добывания пищи хищником при низких значениях плотности жертв. Это, в частности, имеет место при пятнистом размещении жертвы, на которое хищники отвечают также образованием агрегации, а некоторым жертвам в результате этого удается получить частичное убежище ( разд. Кроме того, это можно наблюдать в случаях, когда жертвы имеют настоящие убежища, в которых часть из них спасается от хищников. [12]

Бывает, однако, что соответствие между агрегирующим ответом и распределением поражающего воздействия не всегда соблюдается. Время обработки у данного паразитоида, например, столь велико, что они могут атаковать только очень небольшую часть животных-хозяев в пятнах с высокой плотностью, или в этих пятнах может быть более высокая частота встреч с уже зараженными хозяевами, что может вызвать преждевременный уход паразитоидов из них. Даже в этих случаях популяция жертв имеет частичное убежище. [14]

Страницы: 1 2