Увеличение - плотность - популяция
Cтраница 2
Это мнение увязывается с утверждением Вар-ли, что только простые незапаздывающие факторы, соответствующие регулирующему механизму типа Ферхюльста - Пирла, действительно подходят под концепцию зависимости от плотности, а также с мнением Соломона, что мы не можем исключить паразитов, хищников и других естественных врагов, указанных самим автором этого термина в числе основных примеров в категории зависимости от плотности только потому, что они не непрерывно и не сразу же подавляют увеличение плотности популяции жертвы ( хозяина) согласно точной линейной зависимости. [16]
Неблагоприятное влияние информации отмечено также при скученном содержании крупного рогатого скота, свиней, овец, кур. Увеличение плотности популяций сопровождается возрастанием интенсивности потока информации на каждую особь. Избыток информации приводит к стрессу, задержке роста и развития животных. Нарушается развитие зародышей в утробе беременных коров, овец и свиней. Воспроизводительная функция популяции подавляется. [17]
С увеличением плотности популяции доля паразитов, достигающих половозрелости, снижается, а что еще важнее - стадия, на которой образуются яйца, наступает значительно позже. Это является непосредственной причиной снижения потенциальной скорости роста популяции двуустки. [18]
В результате конкуренции снижается рост и замедляется развитие растений, некоторые из них вымирают. Повышенная гибель, связанная с увеличением плотности популяций, отмечена у моркови, свеклы и других видов культивируемых растений. [19]
Вероятно, эколого-физиологическая разнокачественность генераций как в сезонном, так и в многолетнем аспектах представляет собой достаточно распространенное явление, однако по другим группам животных имеются лишь фрагментарные и не всегда достаточно объективные материалы. У гуппи Poecilia reticulata при увеличении плотности популяции снижается плодовитость и замедляется созревание гонад, что связывают с изменениями генетического состава популяции ( В. [20]
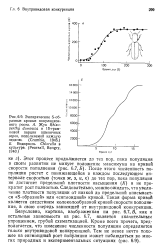
После этого численность популяции растет с понижающейся в каждом последующем интервале скоростью ( точки ж, з, и, к) до тех пор, пока популяция не достигнет предельной плотности насыщения ( / С) и не прекратит рост полностью. Следовательно, можно ожидать, что увеличение плотности популяции от низкой до предельной описывается S-образной или сигмоидной кривой. Такая форма кривой является следствием колоколообразной кривой скорости пополнения, в свою очередь зависящей от внутривидовой конкуренции. Безусловно, картина, изображенная на рис. 6.7, Б, как и остальные зависимости на рис. 6.7, является значительным упрощением, грубой схематизацией. Кроме всего прочего, предполагается, что изменение численности популяции определяется только внутривидовой конкуренцией. [22]
Таким образом, оценка смертности в те периоды, когда она наблюдается, требует введения понятия о компенсационной или распределительной смертности. Так как в действии естественного врага часто наблюдается заметная эластичность и повышение его эффективности связано с увеличением плотности популяции хозяина, отсутствие смертности в какой-то момент в период роста популяции хозяина часто обеспечивает большую смертность, обусловливаемую этим же inn каким-либо другим естественным врагом и последующее время. [23]
Для решения этой задачи использовались простые математические модели - формулы, показывающие наиболее вероятные связи между отдельными, характеризующими состояние популяции величинами: рождаемостью, смертностью, скоростью роста, плотностью ( числом особей на единицу пространства) и др. Математические модели позволяли проверять следствия разных допущений, выявив необходимые и достаточные условия для реализации того или иного варианта популяционной динамики. Перль ( 1879 - 1940) выдвинул так называемую логистическую модель популяционного роста, предполагающую, что по мере увеличения плотности популяции скорость ее роста снижается, становясь равной нулю при достижении некоторой предельной плотности. [24]
Взаимная интерференция может также возникать в тех случаях, когда увеличение плотности консументов ведет к повышенной скорости эмиграции ( рис. 9.11) или когда консументы крадут друг у друга пищу ( как делают многие чайки), или когда жертвы реагируют на присутствие хищника и становятся менее уязвимыми. Однако во всех этих случаях эффект по сути один и тот же: в присутствии других консументов средняя скорость потребления пищи каждой особью понижается и с увеличением плотности популяции хищников этот эффект растет. [25]
Так как многие паразиты имеют два или больше поколения на протяжении жизни одного поколения хозяина, их количественное увеличение может быть весьма существенным. Хотя число жертв одной особи паразита процентно не увеличивается с ростом плотности популяции хозяина, тем не менее общий эффект от популяции паразитов ( если это эффективный паразит при преобладающих условиях) выразится в увеличении процента зараженных жертв на одного паразита с увеличением плотности популяции хозяина. В тех случаях, когда количественная реакция бывает непосредственной, ограничение для такой реакции будет по меньшей мере очень резким у специфичных по хозяевам энтомопаразитов. Следовательно, нет никакого ограничения для процентного увеличения числа зараженных жертв на одного паразита при увеличении плотности популяции хозяина, когда речь идет о популяции паразита, даже если имеется предел эффективной деятельности отдельных особей паразита. [26]
На самом деле асимметричная внутривидовая конкуренция почти без сомнения является общим правилом. Скошенные распределения, которые были показаны, - это одно из возможных его проявлений, но существует и много других Например, Ру-бенштейн ( Rubenstein, 1981), изучая конкуренцию в популяциях черных окуньков-элассом ( Elassoma evergladei), обнаружил, что увеличение плотности популяции приводило к заметному снижению скорости роста, плодовитости самок и репродуктивной активности самцов, по крайней мере это имело место, если сравнивали средних особей, выросших при высокой и низкой плотности. [28]
Реакция у культурных растений разных видов на увеличение плотности популяций неодинакова. Так, при переуплотнении популяции кукурузы вначале отмечается увеличение фитомассы, а затем, начиная с определенной пороговой плотности, урожай этой культуры резко снижается. Увеличение плотности популяций клевера, пшеницы, ячменя приводит к уменьшению облиственности растений. [29]
При первом типе ( кривая 7) скорость роста популяции уменьшается по мере увеличения плотности. Это широко распространенное явление позволяет понять, почему популяции некоторых животных относительно устойчивы. Во-первых, при увеличении плотности популяции наблюдается снижение рождаемости. Так, в популяции большой синицы при плотности меньше одной пары на 1 га на одно гнездо приходится 14 птенцов; когда же плотность достигает 18 пар на 1 га, выводок составляет менее 8 птенцов. Во-вторых, при увеличении плотности популяции меняется возраст наступления половой зрелости. Например, африканский слон в зависимости от плотности популяции может достигать половой зрелости в возрасте от 12 до 18 лет. Кроме того, этот вид при низкой плотности дает приплод 1 слоненок за 4 года, тогда как при высокой - рождаемость составляет 1 слоненок за 7 лет. [30]
Страницы: 1 2 3