Регуляторное воздействие
Cтраница 1
Регуляторные воздействия на биосинтез белка реализуются, по-видимому, на всех его стадиях. Регуляции подлежит работа полимераз, аминоацил - тРНК - синтетазы и рибосом. Воздействия химических соединений на биосинтез далеко не сводятся к мутагенезу. Так, антибиотики влияют на трансляцию кода, воздействуя на рибосомы. Этот антибиотик внедряется в 30S субъединицы рибосомы. [1]
Регуляторные воздействия на всасывание глюкозы проявляются и в действии физиологически активных веществ на различные механизмы ее транспорта, включая движение ворсинок, активность переносчиков и внутриклеточного метаболизма, проницаемость, уровень местного кровотока. [2]
Мишенью различных регуляторных воздействий является ориджин, поэтому для контроля репликации необходимо подавить инициацию в прочих местах. [3]
Мишенью различных регуляторных воздействий является сриджин, поэтому для контроля репликации необходимо подавить инициацию в прочих местах. [4]
В иерархии регуляторных воздействий наиболее сложный механизм, контролирующий количество ферментов в клетке, связан с процессом транскрипции. В случае индукции образованная иРНК участвует в определенной последовательности реакций, называемой трансляцией и заканчивающейся синтезом полипептидных цепей. Регуляция белкового синтеза на уровне трансляции может осуществляться на любом из ее этапов, например на этапе инициации, элонгации и др. Не исключена также возможность изменения времени жизни иРНК под воздействием разных эффекторов, в том числе конечных продуктов метаболических путей. Хотя механизмы регуляции синтеза белка на уровне трансляции еще точно не установлены, ясно, что на этом этапе имеются широкие возможности для регуляции скорости синтеза различных белков. [5]
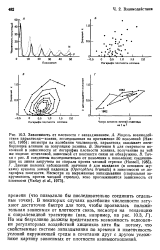
Но мы безусловно должны представлять возможность недооценки регуляторных воздействий хищников хотя бы потому, что свойственные системе запаздывания по времени и изменчивость условий окружающей среды в сочетании друг с другом усложняют картину зависимых от плотности взаимоотношений. [7]
При социализме коренным образом меняются цели, характер, содержание, формы регуляторных воздействий. Это обусловливается рядом объективных причин. Прежде всего социализм органически сочетает разветвленную систему обществ, разделения и кооперации труда с высоким уровнем обобществления средств произ-ва. Это позволяет сознательно координировать работу не только в пределах отд. [8]
В интегрированном состоянии фаговая ДНК реплицируется вместе с бактериальной и подвержена тем же регуляторным воздействиям, что и удвоение бактериальных хромосом. Информация, содержащаяся в фаговой ДНК, в это время не проявляется. Только в результате перехода профага в вегетативное состояние восстанавливается автономия фаговой ДНК и начинается размножение фага. Исключение фаговой ДНК из бактериальной хромосомы происходит, вероятно, путем обращения процессов, приведших к ее включению, и осуществляется очень точно: более 99 % фаговых частиц, освобождающихся из лизогенных клеток, идентичны с исходным ( инфицирующим) фагом. Это означает, что фаговая ДНК при ее выключении выщепляется точно в том же месте, где происходила интеграция. Только в редких случаях ( одном из 100000) выключение ДНК фага происходит аномально ( см. разд. [9]
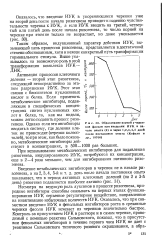 |
Образование корней у черенков фасоли при введении ИУК в начале опыта ( 0 и через 1 2 3 4 5 дней после постановки опыта ( Кефели и др., 19706. [10] |
Несмотря на ведущую роль ауксинов в процессе ризогенеза, природные ингибиторы способны оказывать на первых этапах функционирования ИУК серьезное регуляторное воздействие, направленное на инактивацию избыточных количеств ИУК. Оказалось, что при совместном введении ИУК и фенольных ингибиторов роста в срезанные черенки фасоли ИУК исчезает из черенков значительно быстрее. [11]
Комплементарное спаривание PHKI с РНКП так меняет вторичную структуру последней, что она не может гибридизоваться с областью инициации синтеза ДНК - Интересно, что участок регуляторного воздействия, спаривания РНКП с PHKI отстоит на несколько сотен нуклеотидов от того места, где происходит регулируемое событие - гибридизация РНК с ДНК - Как и в других аналогичных случаях ( например, при регуляции активности генов биосинтеза аминокислот, см. гл. VII), дело здесь в том, что РНКП может в принципе образовать два различных, причем взаимоисключающих варианта вторичной структуры. Лишь один из вариантов способен гибридизоваться с ориджином, а спаривание с PHKI стабилизирует другой вариант и тем самым исключает репликацию. [12]
Комплементарное спаривание PHKI с РНКП так меняет вторичную структуру последней, что она не может гибридизоваться с областью инициации синтеза ДНК - Интересно, что участок регуляторного воздействия, спаривания РНКН с PHKI отстоит на несколько сотен нуклеотидов от того места, где происходит регулируемое событие - гибридизация РНК с ДНК - Как и в других аналогичных случаях ( например, при регуляции активности генов биосинтеза аминокислот, см. гл. VII), дело здесь в том, что РНКП может в принципе образовать два различных, причем взаимоисключающих варианта вторичной структуры. Лишь один из вариантов способен гибридизоваться с ориджином, а спаривание с PHKI стабилизирует другой вариант и тем самым исключает репликацию. [13]
Макс, скорости чтения мРНК рибосомами в отсутствие всяких ограничивающих факторов у прокариот и эукариот составляют соотв. С); регуляторные воздействия могут уменьшить ее до 3 - 10 нуклеотидов в секунду. [14]
Комплементарное спаривание PHKI с РНКП так меняет вторичную структуру последней, что она не может гибридизоваться с областью инициации синтеза ДНК. Интересно, что участок регуляторного воздействия, спаривания РНКП с PHKI отстоит на несколько сотен нуклеотидов от того места, где происходит регулируемое событие - гибридизация РНК с ДНК - Как и в других аналогичных случаях ( например, при регуляции активности генов биосинтеза аминокислот, см. гл. VII), дело здесь в том, что РНКП может в принципе образовать два различных, причем взаимоисключающих варианта вторичной структуры. Лишь один из вариантов способен гибридизоваться с ориджином, а спаривание с PHKI стабилизирует другой вариант и тем самым исключает репликацию. [15]
Страницы: 1 2