Участка - белковая молекула
Cтраница 1
Участки белковой молекулы, которые не относятся к спиральным или складчатым структурам, называются неупорядоченными. При графическом изображении спиральные участки изображают цилиндром, а складчатые структуры - стрелкой. Выделяют понятие надвторичной структуры, представляющей собой закономерное чередование а-спиральных участков и ( J-структур. [1]
Домен - относительно автономные в структурном и функциональном отношении участки белковой молекулы, как правило свернутые в глобулы. [2]
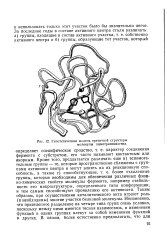 |
Гипотетическая модель третичной структуры молекулы химотрипсиногена. [3] |
Таким образом, при осуществлении каталитического акта играют роль ( и необходимы) многие участки белковой молекулы. Несомненно, что приведенное разделение на четыре типа групп очень условно, поскольку между ними имеется тесная взаимосвязь, и изменения функций в одних группах влекут за собой изменения функций в других. [4]
 |
Водородные связи между парами гетероциклов аденин-тимин и гуанин-цитозин. [5] |
Образование водородных связей с соответствующими белками является одним из основных факторов, обеспечивающих специфическое комплексообразова-ние низкомолекулярных соединений с определенными участками белковых молекул. [6]
Кроме того, высокая способность к образованию водородных связей, наряду с достаточно протяженной цикло-полиэфирной цепочкой, открывает возможность блокировать некоторые хорошо комплементирующие участки белковых молекул и молекул нуклеиновых кислот. [7]
Такая структура, напоминающая слоеный пирог, стабилизирована электростатическими взаимодействиями между ионоген-яым и группами белков, - полиамфолитов, и ионогенными группами липидов, а также гидрофобными взаимодействиями углеводородных частей липидных молекул друг с другом и с неполярными участками белковых молекул. [8]
Полагают [16], что структурные факторы, ответственные за горький вкус, аналогичны по своей природе протонодонорно-протоноакцепторной паре, обусловливающей вкус сладких соединений, за тем исключением, что расстояние между обоими центрами горькой молекулы должно быть равно 0 15 нм. Здесь тоже допускается, что протонодонорный и протоноакцепторный участки белковых молекул, находящиеся на поверхности вкусового окончания, ведут себя как рецептор и образуют водородные связи с горькой молекулой. [9]
Здесь важно отметить, что синтез белка на матрицах нуклеиновых кислот происходит непрерывно, пока клетка жива, и совершается как сложнейший кинетический процесс непрерывного взаимодействия ансамбля переходных состояний. Одни переходные состояния в ансамбле превращаются в конечные продукты - растущие участки белковой молекулы, тогда как другие переходные состояния возникают синхронно ( одновременно) с гибелью первых. По-видимому, в этом состоит поддержание жизни. В построении переходных состояний реакций аминокислоты нуклеиновая кислота - - белок ( ассоциат молекулы белка с нуклеиновой кислотой) участвуют ферменты, витамины, АТФ, вода, многие соединения металлов и неметаллов периодической системы. [10]
Наличие дикетопиперазиновых колец в белковой молекуле, обнаруженное Зелински м, не сразу стало общепризнанным, так как дикетопиперазины могли образоваться из свободных аминокислот и таким образом не быть одним из продуктов гидролитического распада белковой молекулы, а могли оказаться вторичным образованием, возникающим после того, как молекула белка распалась на отдельные аминокислоты. Однако Зелинский и его сотрудники рядом тщательных исследований доказали, что ди-кетопиперазиновые кольца действительно являются участками белковой молекулы. [11]
Образование такой детерминанты способствует восприятию антигенраепознающими Т - лимфоцитами молекулы модифицированного белка как чужого. По-видимому, не обязательно гаптен будет модифицировать запрещенную детерминанту белка, к которой имеется естественная толерантность. Новая детерминанта может возникнуть на любом участке белковой молекулы. Большинство химических аллергенов не присоединяется к участкам молекулы белка, которые распознаются как специфическая детерминанта, ибо в противном случае чужеродные белковые носители искусственных конъюга-тов не служили бы индуктором иммунного ответа к носителю, а антитела бы против них не реагировали с интактными аллогенными белками. [12]
Кроме того, в этой главе рассматриваются три типа бифункциональных реагентов. При обработке такими реагентами белков образуются внутри - и межмолекулярные связи. Реагенты второго типа имеют аналогичную группировку R, а также группу А, обладающую специфическим сродством к боковой цепи одной из аминокислот. Благодаря этой группе реагент, сорбируется на определенном участке, например в области связывания, узнавания или на регуляторном участке фермента, а также в областях с иными свойствами, например, на гидрофобных, основных или кислотных, положительно или отрицательно заряженных областях или участках белковой молекулы. Реагенты третьего типа обозначаются S-О - А или S-О - R, где S означает сигнальную группировку. При сорбции или химическом связывании такой группировки с участком белковой молекулы она ( благодаря изменению окраски, флуоресценции или спинового состояния) передает информацию о своем непосредственном окружении. [13]
Страницы: 1