Участка - белок
Cтраница 1
 |
Распределение липидов между наружной ( а и внутренней ( б сторонами бислоя в мембранах эритроцитов ( I, вируса гриппа ( II и саркоплазматического ретикулума кролика ( III. [1] |
Участки белка, которые обращены во внеклеточную среду, могут подвергаться гликозилированию. В мембранах растений и бактерий полисахара играют самостоятельную роль, образуя наружную оболочку. [2]
Сигмоидный характер ской кривой аллостерического фермента объясняют кооперативным взаимодействием между связывающими участками белка. [3]
Первоначально на основании анализа профиля гидрофобности полнпептидиых цепей холинорецептора было предположено, что каждая субъединица имеет по четыре трансмембранных фрагмента, a N - и С-концевые участки белков расположены снаружи постси-наптической мембраны. Однако с помощью антител против синтетических пептидов, соответствующих С-концевым областям различных субъединиц, удалось доказать локализацию С-концевого фрагмента на цитоплазматической стороие мембраны, вследствие чего субъединицы холинорецептора должны иметь по крайней мере по пять трансмембранных сегментов. [4]
 |
Возможные способы образования водородных связей в трипептидах. [5] |
Свернутые формы интересны постольку, поскольку они преобладают в неполярных растворителях, и, кроме того, некоторые из них могут встречаться в олигопептидах, полипептидах и нерегулярных участках белков. [6]
Небольшие отличия в последовательностях аминокислот наблюдаются на поверхностных участках белка. Некоторые же последовательности аминокислот характерны для всех образцов цитохрома с. Это показывает, что мутации, затрагивающие эти участки, всегда летальны. Это показывает, что в основном функция цитохрома с осталась практически неизменной в процессе эволюции, продолжавшейся миллиарды лет. [7]
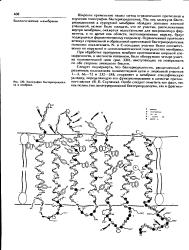 |
Топография бактериородопсина в мембране. [8] |
Так как молекула бакте-риородопсина в пурпурной мембране обладает довольно жесткой упаковкой, можно было ожидать, что ее участки, расположенные внутри мембраны, окажутся недоступными для макромолекул ферментов, в то время как области, экспонироваииые наружу, будут подвергаться ферментативному гидролизу. Ограниченный протеолнэ везикул с правильной и обращенной ориентацией бактериородопсина позволил локализовать N - и С-концевые участки белка соответственно на наружной и цитоплазматической поверхностях мембраны. [9]
В модельном Са-пептидном комплексе [292] длины связей Са-О составляют 220 - 260 пм. Эти данные объясняют преимущественное содержание иона Са ( П) внутри окружения карбонильных и карбоксильных донорных атомов кислорода и показывают, что ионы Са ( П) могут легко размещаться в неправильном координационном окружении. С точки зрения предположения Уильямса [290] о Са ( П) как мостике его роль в альбумине карпа может заключаться в стабилизации структуры связыванием карбоксильных групп боковых цепей аминокислот между а-спиральными участками белка. [10]
ДНК, к которому этот белок присоединяется. В работе [137] предполагается, что участок регуляторного белка состоит из двух антипараллельных сегментов полипептидной цепи, образующих ( - структуру. Узнавание основано на компле-ментарности этой структуры и последовательности пар оснований ДНК - Важное свойство такой последовательности состоит в асимметричном распределении гуанинов между двумя нитями ДНК - В предлагаемом коде шесть основных аминокислотных остатков ( Сер, Тре, Асп, Гис, Глн, Цис) и их последовательность в стереоспецифичном участке белка определяет последовательность пар оснований, с которой данный белок преимущественно связывается. Код, разработанный на основе стереохимии, подтвержден на примере взаимодействия Лак-репрес-сора с Лак-оператором ( см. стр. [11]
 |
Схема оперона. [12] |
Предполагается, что участок регуляторного белка состоит из двух антипараллельных сегментов полипептидной цепи, образующих - структуру. Узнавание основано на комплемен-тарности этой структуры и последовательности нуклеотидных пар ДНК. Важное свойство такой последовательности состоит в асимметричном распределении гуанинов между двумя нитями ДНК. В предлагаемом коде шесть аминокислотных остатков - Сер, Тре, Асп, Гис, Глн, Цис - и их последовательность в сте-реоспецифичном участке белка определяет последовательность пар оснований ДНК, с которой данный белок преимущественно связывается. Код, разработанный на основе стереохимии, подтвержден взаимодействием Лак-репрессора с Лак-опероном и другими примерами. [13]
Причины специфичности взаимодействия Cibacron Blue с белками остаются неясными. Первоначально этот лнганд был успешно использован для очистки ряда дегидрогеназ, киназ и мутаз - ферментов, имеющих в активном центре пространственную структуру так называемой динуклеотидной складки, образованной примерно 120 аминокислотами. В этой структуре связываются НАД, ADP или АТР. Было замечено, что пространственная конфигурация красителя напоминает таковую для НАД, причем сульфогруппы расположены примерно в тех же местах, что фосфаты у кофермента. Поэтому Ион предлагает отнести Cibacron Blue к числу имфилитов - сорбентов со смешанной ионно-гидрофобной функцией, для которых некая биоспецифичность получается в результате случайного пространственного соответствия комбинации этих функций какому-то участку белка. Вместе с тем отмечалось, что краситель Procion Red HE3B имеет большее сродство к НАДФ - зависимым дегидрогеназам и киназам, но меньшее - к НАД - зависимым, чем Cibacron Blue. [14]
Реакция аминогруппы с фенилизотиоцианатом нашла широкое применение для определения N-концевых аминокислот белка или пептидов. Впервые этот метод был предложен в 1950 г. Эдманом. При взаимодействии фенилизотиоциана-та с а-аминогруппами N-концевых аминокислот белка образуется фенилтиокарбамильное производное белка. При обработке фенилтиокарбамильного производного белка кислотой происходят циклизация и отщепление N-концевых остатков в виде фенилтиогидантоинов. Последние могут быть хроматографически разделены и идентифицированы; можно расщепить их и до свободных аминокислот и определить последние. Остающийся после отщепления фенилтиогидантоина белок содержит на одну аминокислоту меньше, чем исходный. Таким образом можно постепенно, отщепляя одну аминокислоту за другой, определить их последовательность в N-кон-цевом участке белка. [15]
Страницы: 1