Учет - кулоновское взаимодействие
Cтраница 2
В последующих многочисленных работах [32-35] количественные оценки, сделанные Де Во и Тиноко, неоднократно критиковались, в частности все авторы указывали на недопустимость приближения точечных диполей и проводили учет кулоновских взаимодействий, центрируя заряды на атомах оснований. Корректность той или иной параметризации для различных составляющих свободной энергии полинуклеотида и в настоящее время не может быть доказана с определенностью. Между тем основные идеи были высказаны именно Де Во и Тиноко и ими же теоретически было установлено наличие сильных стэкинг-взаимодей-ствий, стабилизирующих структуру одиотяжевых и двухтяжевых полинуклеотндов, в то время как водородные связи, на которых основывалась модель Уотсона и Крика, играют второстепенную роль. Анализируя результаты расчетов различных авторов, можно отметить, что хотя между ними нет количественного согласия, качественный вывод Де Во и Тиноко о доминирующей роли стэ-кинг-взаимодействнй в стабилизации конформации дикулеозид-фосфатов, олигомеров, а также однотяжевых и двухтяжевых по-линуклеотидов разделяется всеми авторами. [16]
Поэтому расчеты по моделям ( 179) и ( 183) могут определить масштаб влияния различных предположений на матрицу корреляций флуктуации и рассчитываемые с их помощью кинетические величины. Учет кулоновского взаимодействия между связанными и свободными электронами уменьшает матрицу корреляций. Учет связи флуктуации увеличивает матрицу корреляций, уменьшая величину флуктуации чисел заполнения связанных состояний. Выбор ансамбля является независимым приближением ячеечной модели неидеальной плазмы, в которой само разделение электронов на связанные и свободные строго не определено. [17]
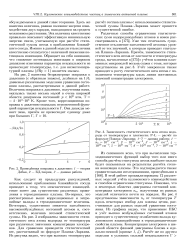
Из сказанного ясно, что при вычислении термодинамических функций выбор того или иного способа расчета статсуммы атома наиболее сильно будет сказываться на результатах расчета степени ионизации плазмы. В этой работе проанализированы 12 различных моделей учета кулоновского взаимодействия и способов ограничения статсуммы. Показано, что в некоторых областях диаграммы состояний концентрация электронов пе, полученная из разных моделей отличается почти на порядок. На рис. 4 представлены зависимости пе от температуры Т вдоль некоторых изобар для плазмы цезия, рассчитанные для разных моделей уравнения состояния. Из графика видно, что применение ПБС и учет высоко возбужденных состояний атомов приводит к существенному ослаблению вклада кулоновского взаимодействия в термодинамические свойства плазмы. [19]
 |
Многоканальный рассеиватель S. Падающая слева в канале j волна. [20] |
Обобщение на случай конечных частот ( Buttiker, 1993) и учет кулоновского взаимодействия также являются очень важными. [21]
На практике чаще всего возникает задача формирования потоков заряженных частиц, а вычисление отдельных траекторий требуется редко. Хотя эти траектории и впредь будут играть определенную роль, необходимо познакомиться с описанием свойств коллективов частиц. Отложим на время учет кулоновского взаимодействия между частицами самого потока, т.е. будем считать плотность заряда пренебрежимо малой. [22]
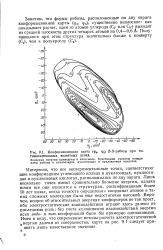 |
Конформационная карта ( ф4, р5 P-D - рибозы при оптимизированных валентных углах. [23] |
Интересно, что все экспериментальные точки, соответствующие конформациям углеводного кольца в нуклеозидах, нуклеотидах и нуклеиновых кислотах, расположились по дну оврага. Лишь несколько точек имеют сравнительно большие энергии, однако почти все они относятся к структурам, расшифрованным более 10 лет назад, когда точность определения координат атомов в таких сложных молекулах была еще недостаточно высокой. Впрочем, вопрос об относительных энергиях конформеров не так прост, ибо электростатические взаимодействия играют здесь далеко не последнюю роль: без учета кулоновских взаимодействий энергии всех эндо - и э / сзо-конформаций были бы примерно одинаковыми. [24]
Мы видели, что теория Келли - Фаликова может, по-видимому, объяснить качественно различные аномальные и необъясненные явления на поверхностях ( 111) и ( 110) и иа поверхности ( 100) в условиях деформации. Однако наряду с этими достижениями имеется серьезная проблема, связанная с абсолютными величинами параметров модельного гамильтониана. Келли и Фаликов [923, 924] пытались оценить величину У, используя трехмерный экранированный кулоновский потенциал, в котором роль подгоночного параметра играет радиус экранирования. Возможны другие способы оценки V на основе более сложных методов учета электрон-электронных кулоновских взаимодействий, рассмотренных в § 6 гл. Один из таких способов опирается на выражение для эффективного g - фактора, которое может быть получено из гамильтониана (7.50) в приближе - нии Хартри - Фока. [25]
Простые вычисления показывают, что процесс, описываемый уравнением ( 50), термодинамически вероятен для ферментативной реакции NADH с ацетальдегидом. Потенциал восстановления ацетальдегида в диметилформ-амиде равен - 2 25 в [229], а в специфической среде у активного центра дегидро-геназы, вероятно, меньше. Если предположить, что для образования протонированной формы NADH -, генерируемой электронным переходом, требуется еще 0 1 в, то для реакции, приведенной на схеме ( 54), будет необходимо не более 3 19 в или 73 5 ккал / моль без учета кулоновского взаимодействия и других специфических взаимодействий фермента. Кулоновское взаимодействие описывается уравнением Е - - 330 qiqzIDr и на расстоянии 3 0 А равно 56 ккал / моль, а в переходном состоянии для расстояния между реагентами 2 5 А составляет 66 ккал / моль. [26]
Затем будет выяснено влияние сил связи, действующих на электроны, и получена классическая формула Бора для потерь энергии. Далее обсуждаются ее квантовые модификации, затем учитывается влияние поляризации среды, рассматриваются потери энергии в электронной плазме. Вслед за этим будет исследовано упругое рассеяние частиц на ядрах и многократное рассеяние. В конце рассматривается вопрос об электропроводности плазмы - с учетом экранированного кулоновского взаимодействия. [27]
Страницы: 1 2