Белковый фактор - инициация
Cтраница 1
 |
Схематическое изображение взаимодействия формилметионил-т РНК и мРНК с 30S субчастицей рибосомы ( а и транслирующей ( функционально активной 70S рибосомой ( б. [1] |
Аналогичные белковые факторы инициации обнаружены также в эука-риотических клетках. Они получены в чистом виде: eIF - 2 состоит из а -, 3 - и у-субъединиц ( мол. Укажем также, что в синтезе белка их роль тождественна роли инициаторных белков у прокариот. Отличительной особенностью синтеза белка у эукариот является, кроме того, наличие среди 10 белковых факторов инициации еще одного белка, названного кэп-связы-вающим. Соединяясь с 5 -участком кэп мРНК, этот белок содействует образованию комплекса между мРНК и 40S рибосомной субчастицей. Необходимо отметить, что до сих пор не раскрыты тонкие молекулярные механизмы участия белковых факторов инициации как у про -, так и у эукариот в сложном процессе синтеза белка. [2]
К настоящему времени выяснена природа белковых факторов инициации. Все они получены в высокоочищенном состоянии с примерными молекулярными массами 9000, 10000 и 22000 соответственно. IF-3 обеспечивает узнавание участка на молекуле мРНК, к которому присоединяется формилметионил-т РНК. Белковый фактор IF-2, вероятнее всего, способствует объединению 30S и 50S субчастиц после того, как на первой субчастице уже присутствуют инициирующие кодоны мРНК, N-формилметионил-т РНК, IF-3, IF-1 и ГТФ. [3]
Стадия инициации, являющаяся точкой отсчета начала синтеза белка, требует соблюдения ряда условий, в частности наличия в системе, помимо 70S ( или 80S) рибосом, инициаторной амино-ацил - тРНК ( аа-т РНК), инициирующих кодонов в составе мРНК и белковых факторов инициации. Экспериментально доказано, что синтез белка инициирует единственная аминокислота - метионин. Соответственно эти тРНК принято обозначать тРНК Мет и тРНК ет. Укажем также, что эукариотическая клетка не нуждается в формилировании метионина. [4]
 |
Схематическое изображение взаимодействия формилметионил-т РНК и мРНК с 30S субчастицей рибосомы ( а и транслирующей ( функционально активной 70S рибосомой ( б. [5] |
Как только мРНК присоединяется к комплексу, высвобождается белковый фактор IF-3 и оставшийся комплекс легко присоединяет 50S субчастицу, образуя транслирующую, т.е. функционально активную, 70S рибосому. В процессе этих перестроек рибосомы освобождают остальные белковые факторы инициации и продукты гидролиза ГТФ ( ГДФ и неорганический фосфат), энергия которого расходуется, по-видимому, на формирование инициирующего 70S комплекса рибосомы. В этом комплексе формилметионил-т РНК оказывается прикрепленной к пептидилсвязывающему центру рибосомы. [6]
 |
Рибосомы про - и эукариот.| Полная рибосома и компоненты инициации трансляции. [7] |
Инициация начинается с образования малых инициирующих комплексов, которые затем ассоциируют в большой инициирующий комплекс. В этом процессе участвуют рибосомы, мРНК, амино-ацил - тРНК, белковые факторы инициации: ( IF, IF2 и IF3), а также макроэрг ГТФ. [8]
Репликация ДНК представляет полуконсервативный процесс, в котором дочерние клетки получают по одной нити материнской и одной нити вновь синтезированной ДНК. Репликация хромосомы начинается в участке, называемом oriC ( от англ, origin), присоединением белковых факторов инициации. Это сопровождается расплетением двойной спирали ДНК с образованием двух репликативных вилок, на каждой из которых начинается встречный синтез второй нити. Для синтеза существует сложный ферментный аппарат; обеспечивающий расплетение нитей ( ДНК-гиг раза или топоизомераза), хеликаза, праймаза с праймером РНК, ДНК-полимераза I и ДНК-полимераза III. Для ее функционирования необходимо поступление предшественников в виде праймеров РНК, в свою очередь образуемых из пентоз, оснований, АТФ - из цитоплазмы и ферментов -: изрибосомального аппарата. [9]
Менее определена функция группы регуляторных белков и пептидов, поскольку, в известной степени, эту роль выполняют любые белки. Сюда относят белково-пептидные вещества, не вошедшие в состав вышеупомянутых групп, но весьма важные для функционирования отдельных звеньев клеточного механизма, например гистоны и репрессоры, регулирующие активность генов, воротные белки мембранных каналов, рибосома л ьные белковые факторы инициации и элонгации ( см. с. [10]
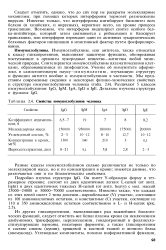 |
Свойства иммуноглобулинов человека. [11] |
Следует отметить, однако, что до сих пор не раскрыты молекулярные механизмы, при помощи которых интерфероны тормозят размножение вирусов. Известно только, что интерфероны ингибируют биосинтез всех белков ( и хозяйских, и вирусных), вероятнее всего, на уровне процесса трансляции. Возможно, что интерферон индуцирует синтез особого белка-ингибитора, который затем связывается с рибосомами и блокирует трансляцию, или интерферон переводит один из активных эукариотических белковых факторов инициации в неактивный фактор путем фосфорилиро-вания. [12]
Аналогичные белковые факторы инициации обнаружены также в эука-риотических клетках. Они получены в чистом виде: eIF - 2 состоит из а -, 3 - и у-субъединиц ( мол. Укажем также, что в синтезе белка их роль тождественна роли инициаторных белков у прокариот. Отличительной особенностью синтеза белка у эукариот является, кроме того, наличие среди 10 белковых факторов инициации еще одного белка, названного кэп-связы-вающим. Соединяясь с 5 -участком кэп мРНК, этот белок содействует образованию комплекса между мРНК и 40S рибосомной субчастицей. Необходимо отметить, что до сих пор не раскрыты тонкие молекулярные механизмы участия белковых факторов инициации как у про -, так и у эукариот в сложном процессе синтеза белка. [13]
Аналогичные белковые факторы инициации обнаружены также в эука-риотических клетках. Они получены в чистом виде: eIF - 2 состоит из а -, 3 - и у-субъединиц ( мол. Укажем также, что в синтезе белка их роль тождественна роли инициаторных белков у прокариот. Отличительной особенностью синтеза белка у эукариот является, кроме того, наличие среди 10 белковых факторов инициации еще одного белка, названного кэп-связы-вающим. Соединяясь с 5 -участком кэп мРНК, этот белок содействует образованию комплекса между мРНК и 40S рибосомной субчастицей. Необходимо отметить, что до сих пор не раскрыты тонкие молекулярные механизмы участия белковых факторов инициации как у про -, так и у эукариот в сложном процессе синтеза белка. [14]
Страницы: 1