Форма - кривая - выживание
Cтраница 1
 |
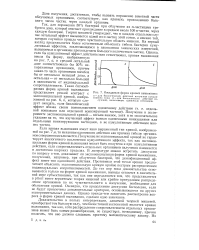
Кривые выживания. 1 - человек или крупные млекопитающие. 2 - гидра или чернохвостый олень. 3 - устрица или рыба. [1] |
Форма кривой выживания часто меняется при изменении плотности популяции. [2]
Такого общего увеличения дозы инактивации с увеличением плотности ионизации и следует ожидать, если мы имеем дело с эффектом облучения, причиной которого являются единичные ионизации ( ср. Вместе с формой кривой выживания и независимостью дозы инактивации от интенсивности облучения факт увеличения этой дозы с увеличением плотности ионизации служит экспериментальным доказательством того, что инактивация вируса представляет собой пример эффекта, вызываемого единичной ионизацией. [3]
Во-первых, следует определить, каким образом происходит возрастание численности организмов или клеток, подвергнутых действию излучения, при увеличении дозы. Если изучается летальное действие излучения, то это определение эквивалентно нахождению формы кривой выживания, при построении которой по оси ординат откладываются относительные количества организмов, оставшихся неповрежденными, а по оси абсцисс - соответствующие им дозы излучения. Во-вторых, нужно выяснить, как зависит вызываемый данной дозой эффект от интенсивности излучения. В-третьих, необходимо определить относительные эффективности различных излучений в зависимости от их типа или длины волны. Последнее особенно важно; именно то обстоятельство, что теория мишеней способна с успехом объяснить зависимость эффекта данной дозы от длины волны или типа излучения, и определяет ее принципиальную ценность. [4]
К третьему классу действий излучений относятся такие эффекты, для возникновения которых необходимо, чтобы внутри мишени было произведено большое количество ионизации и, следовательно, сквозь мишень должно пройти несколько ионизирующих частиц. Чем выше интенсивность ионизации, тем больше ионизации происходит на каждый микрон пути частицы и тем меньше нужно ионизирующих частиц, чтобы вызвать эффект. Очевидно, различие в ионизирующей способности должно проявляться в изменении формы кривой выживания для различных длин волн или типов излучения. [5]
Очень возможно, что, исходя из теории мишеней, можно правильно истолковать и другие действия излучений, но в отношении этих случаев пока еще нет достаточных экспериментальных доказательств, позволяющих рассматривать теорию мишеней как нечто большее, чем рабочая гипотеза. Попытки интерпретировать некоторые биологические действия излучений при помощи теории мишеней без достаточных для этого оснований приводили часто к бесплодной полемике. Поэтому здесь не рассматриваются примеры применения теории мишеней только на основе формы кривой выживания. [6]
 |
Ожидаемая форма кривых выживания. [7] |
Если кривая выживания имеет явно выраженный тип кривой, изображенной на рис. 7 а, то механизм одиночного действия как причина гибели организмов совершенно исключается. Получение же экспоненциальной кривой не гарантирует аналогичного исключения кумулятивного эффекта, так как экспоненциальная форма кривой выживания может быть получена и при кумулятивном действии, если сопротивляемость отдельных организмов излучения изменяется в достаточно широких пределах. В литературе можно встретить дискуссии по вопросу о том, доказывает ли экспоненциальная форма кривой выживания, полученная, например, при облучении бактерий, что дезинфекционный эффект имеет тип одиночного действия. Противники этой точки зрения предпочитают объяснять экспоненциальную кривую крайне пестрым распределением индивидуальных сопротивляемостей. До тех пор пока доказательства основываются только на форме кривой выживания, выводы остаются в значительной мере субъективными, так как они определяются тем, что представляется a priori менее вероятным: теория мишеней или крайне причудливое распределение организмов по их чувствительности к излучению, необходимое для объяснения кривой. Очевидно, что продолжение дискуссии бесполезно, если не будут привлечены дополнительные критерии, основывающиеся на других экспериментальных данных. [8]
Наличие небольших систематических отклонений от экспоненциальной кривой не указывает, однако, на непригодность теории мишеней, если, конечно, не были специально приняты меры для исключения усложняющих явление факторов, способных исказить экспоненциальный характер кривой выживания. Так, если при облучении бактерий в суспензии образуются местные скопления, кривая выживания делается слегка сигмообразной, так как требуется несколько попаданий, каждое в отдельную молекулу, чтобы лишить такое скопление способности к дальнейшему размножению с образованием колоний. Если облучаемые организмы обладают различной устойчивостью к излучению, то более чувствительные будут убиты быстрее, чем более устойчивые, и кривая выживания, построенная в логарифмическом масштабе, будет несколько выпуклой. Если для распространения исследований в область почти полного уничтожения организмов применяемые в опыте дозы доводятся до очень высоких, это может привести к возникновению добавочных летальных факторов ( например, к образованию ядовитых продуктов распада среды), что также может обусловить отклонение кривой от экспоненты. Очень часто подобные осложняющие обстоятельства отсутствуют, но если они есть, то, естественно, ослабляют доказательства, основанные на форме кривой выживания и законности применения теории мишеней к исследуемому явлению. [9]
Препарат этот был облучен в нескольких различных концентрациях, а также в сухом виде. Добавление к разбавленному раствору желатины приводило к повышению дозы инактивации почти до величины, характерной для сухого препарата. Полученные результаты приведены в табл. 30 и иллюстрируются рис. 16, а. Вывод из них сводится к тому, что непрямое действие может быть в значительной мере подавлено, если применять достаточные концентрации белка - все равно, вирусного или желатины. Однако в разбавленных растворах непрямое действие преобладает. Форма кривой выживания не может служить критерием прямого или непрямого действия, так как в обоих случаях получаются экспоненциальные кривые. [10]
 |
Кривые выживаний для некоторых видов животных, представленные в полулогарифмическом масштабе ( по. Пианка, 1981. [11] |
Первый тип ( кривые 7) соответствует ситуации, когда большее 1исло особей имеет одинаковую продолжительность жизни и умира-зт в течение очень короткого отрез-са времени. Кривые характеризуются сильно выпуклой формой. Такие сривые выживания свойственны че-повеку ( рис. 7.3, /), причем кривая выживания для мужчин по сравнению с аналогичной кривой для женщин менее выпуклая, поэтому страховой полис для мужчин в большинстве стран Запада в 1 5 раза дороже, чем для женщин. Для большинства копытных кривые выживания также выпуклые ( рис. 7.4), хотя и в различной степени для разных видов, а также в зависимости от пола. Второй тип ( рис. 7.3, 2 свойствен видам, коэффициент смертности которых остается постоянным на протяжении всей их жизни. Поэтому кривая выживания трансформируется в прямую линию. Такая форма кривой выживания свойственна пресноводной гидре. [12]
С другой стороны, более крупные нехватки, захватывающие свыше 50 дисков хромосомы слюнной железы, не встречаются даже в гетерозиготном состоянии, точно так же, как никогда не встречаются в клетках слюнных желез и дицентрические хромосомы. Однако есть основания думать ( Демерец, Кауфман, Сеттон и Фано, 1941), что делеции происходят так же часто, как и равновеликие инверсии, и что асимметричные обмены, приводящие к возникновению дицентрических и ацентрических хромосом, приблизительно так же часты, как и симметричные обмены, дающие начало моноцентрическим хромосомам. Поэтому следует предположить, что более крупные нехватки ведут себя как доминантные летали. Ацентрическая хромосома, поскольку она лишена центромера, ответственного за начало отхождения дочерних хромосом к полюсу в делящейся клетке, легко может остаться вне обоих дочерних ядер. Два центромера дицентрической хромосомы могут стремиться отойти к противоположным полюсам. Не приходится поэтому сомневаться, что более крупные нехватки и асимметричные обмены ведут себя как доминантные летали. Более того, форма кривой выживания ( см. рис, 27) не согласуется с предположением, что дицентрические хромосомы и крупные делеции целиком ответственны за доминантные летали. Кривые выживания, нанесенные на рис. 27 на логарифмической шкале, в начальной своей части линейны. [13]
Страницы: 1