Полиплоидные формы
Cтраница 1
Полиплоидные формы многих групп растений имеют пониженную фертильность. Это явление наблюдается и у картофеля. Но степень фер-тильности не имеет у картофеля такого значения, как у зерновых культур. Того уровня плодовитости, которым обладают экспериментально полученные полиплоиды, вполне достаточно для проведения с ними селекционной работы, тем более, если принять во внимание их лучшую скрещиваемость с другими видами. [1]
Полиплоидные формы свеклы устойчивы к гербицидам, что позволяет повысить дозировку гербицидов при предпосевных обработках и добиться высокой гибели сорняков без снижения урожая корней. [2]
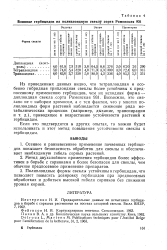 |
Влияние гербицидов на полиплоидную свеклу сорта Рамонская 931. [3] |
Полиплоидные формы свеклы устойчивы к гербицидам, что позволяет повысить дозировку гербицидов при предпосейных обработках и добиться высокой гибели сорняков без снижения урожая корней. [4]
Полиплоидные формы второго года жизни являются позднеспелыми, но в настоящее время после селекционных отборов по раннеспелости на семенниках они также не являются позднеспелыми. [5]
При набухании семян в растворе колхицина были получены полиплоидные формы у табака, райграса, красного перца, разновидностей капусты. В работе с картофелем семена проращивают в чашках Петри на субстрате, состоящем из равных частей 0 5 % - ного колхицина и 2 % - ного агара. После прорастания семена промывают и высаживают. [6]
У растений, способных размножаться листовыми черенками, можно получить полиплоидные формы путем регенерации растений из листьев, обработанных колхицином. [7]
Апомиксис также влечет за собой неспособность к образованию гибридов, и благодаря этому полиплоидные формы действенно изолируются от исходных диплоидных форм. Так как апомиксис широко распространен не только среди животных, но и среди растений, нужно несколько остановиться на его роли в образовании видов. [8]
Однако определение видового статуса в этих случаях чрезвычайно усложняется другим обстоятельством - всеобщим распространением в апомиктических группах полиплоидии. Полиплоидные формы у апомиктов могут быть как авто -, так и аллополиплоидами или же представляют собой смесь этих двух форм. Изменения признаков при этом часто носят количественный характер, что сильно затрудняет работу систематиков, пытающихся разобраться во всем этом многообразии форм. [9]
Из приведенных данных видно, что если тетраплоидные формы доведены до полной стабильности по плоидности, то по энергии корнеоб-разования они превосходят исходные диплоидные формы в течение всего вегетационного периода. Очевидно полиплоидные формы, так же как и дигшоиды, могут быть раннеспелыми или позднеспелыми по одному из показателей: энергии корнеобразования или сахаронакоплению. На данном этапе основная часть тетраплоидных сортов является раннеспелой по энергии корнеобразования. Поэтому относить полиплоидные формы первого года жизни к позднеспелым не следует. [10]
Для того чтобы посев свеклы и обработка почвы гербицидами осуществлялись одновременно, необходимо механизировать этот процесс. Однако имеются сведения, что полиплоидные формы растений более устойчивы к болезням растений и неблагоприятным факторам среды. [11]
Для того чтобы посев свеклы и обработка почвы гербицидами осуществлялись одновременно, необходимо механизировать этот процесс. Однако имеются сведения, что полиплоидные формы растений более устойчивы к болезням растений и неблагоприятным факторам среды. Если это предположение окажется верным для стойкости полиплоидной свеклы к гербицидам, то отпадет необходимость работать над двухслойным применением гербицидов, так как простое предпосевное применение препаратов более удобно для практических работников. [12]
Наследственная изменчивость в природе не ограничивается исключительно рекомбинационной изменчивостью, но усложняется изменениями в структуре и числе хромосом. Что касается числа хромосом, то особый интерес представляют полиплоидные формы. XXV мы показали, как путем скрещивания и удвоения хромосом могут возникнуть новые полиплоидные виды. [13]
Из приведенных данных видно, что если тетраплоидные формы доведены до полной стабильности по плоидности, то по энергии корнеоб-разования они превосходят исходные диплоидные формы в течение всего вегетационного периода. Очевидно полиплоидные формы, так же как и дигшоиды, могут быть раннеспелыми или позднеспелыми по одному из показателей: энергии корнеобразования или сахаронакоплению. На данном этапе основная часть тетраплоидных сортов является раннеспелой по энергии корнеобразования. Поэтому относить полиплоидные формы первого года жизни к позднеспелым не следует. [14]
При обсуждении роли полиплоидии в эволюции растений мы должны учитывать следующие обстоятельства. Полиплоиаизация хромосомного набора вида приводит не только к увеличению дозы соответствующего гена, что само по себе дает значительный фенотипический эффект и может иметь непосредственное адаптивное значение, но при дальнейших изменениях генов в результате мутагенеза полиплоидизация приводит к созданию новой генетической системы. Таким путем могут развиваться, вероятно, комплементарные и полигенные системы, имеющие определенное адаптивное значение. Известно, что каждый генокон - тролируемый процесс осуществляется системой, состоящей из многих генов. Продолжительность периода, в течение которого происходят метаболические процессы, также зависит от числа последовательно активизирующихся генных единиц, и полиплоидия может рассматриваться как один из механизме создания такой системы. Исследования Маккей [ 58 ] показали, что эволюция пшеницы от примитивных ломкоколосых и спель-тоидных типов до прочноколосых и легко обмолачивающихся форм шла именно в направлении развития полигенной системы параллельно с возникновением полиплоидных видов. Кроме того, элементарные наборы хромосом, составляющие кариотип организма, при полиплоидизации или гибридизации не остаются тождественными, поскольку каждая хромосома с течением времени подвергается изменениям, что может и должно приводить к диплоидизации видов. При сохранении основного числа хромосом их генная структура изменяется. Мы хорошо знаем, что естественные полиплоидные формы и виды, прошедшие некоторый эволюционный путь развития, ведут себя функционально как диплоиды. Это последнее непосредственно сказывается на прохождении у них мейоза: как правило, образуются биваленты, а не мультиваленты, которые обычно характерны для только что полученных экспериментальных поли-лпидов. При этом вследствие увеличения числа хромосом при полиплоидии и генетической дифференциации между ранее гомологичными хромосомами будет повышаться индекс рекомбинации и изменчивости, что является источником видообразования. Кроме того, число жизнеспособных рекомбинантов при полиплоидизации возрастает, так как добавление, утеря или замена отдельных хромосом в мейозе происходит у полиплоидов с меньшим нарушением генетического баланса, чем у диплоидов. Такие особи могут быть исходными для образования производных видов. [15]
Страницы: 1