Фосфорилирование - фермент
Cтраница 1
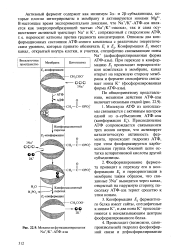 |
Механизм функционирования Na / K - ATO - a3bi. [1] |
Фосфорилирование фермента приводит к переходу его в кон-формацию Е2 и переориентации в мембране таким образом, что связанные 3Na выводятся через канал, открытый на наружную сторону, поскольку АТФ-аза теряет сродство к этим ионам. [2]
Фосфорилирование фермента приводит к переходу его в конфор-мацию Е2 и переориентации в мембране таким образом, что связанные 3Na4 выводятся через канал, открытый на наружную сторону, поскольку АТФ-аза теряет сродство к этим ионам. [3]
 |
Модель активного центра щелочной фосфатазы. а - фосфорилирование фермента. б - перенос фосфата на воду и другие нуклеофилы. [4] |
После фосфорилирования фермента происходит нуклеофильная атака на образовавшееся промежуточное соединение. Нуклео-филом может быть не только вода. Вильсон и др. [37] показали, что ряд акцепторов фосфата, таких, как этаноламин и этилен-гликоль, эффективно конкурирует с водой, даже если их концентрация 1 М или меньше. [5]
Активная протеин-киназа катализирует фосфорилирование ферментов - фосфорилазы и гликогенсинтазы. Образуется активная фосфорилированная форма фосфорилазы а и неактивная фосфорилированная форма гликогенсинтазы D; такие изменения обеспечивают фосфоролити-ческий распад гликогена. [6]
При этом происходит фосфорилирование фермента с отщеплением кислотного остатка X. Природа X не влияет на строение ингибирован-ного фермента, но сильно воздействует на процесс его ингибирования. Очень активными ингибиторами холинэстеразы являются фосфорорганические вещества, несущие заряд в кислотном остатке. Например, усиление действия происходит при окислении атома серы в тиоэфир-ном радикале. [7]
Конечным результатом этих реакций является в первом случае фосфорилирование фермента, во втором - фосфорилирование гид-роксила. [8]
Реакция фосфорилирования ( 2 - 24) дает дополнительные, хотя и косвенные доказательства участия имидазольной группы в ацилировании или фосфорилировании фермента. Химотрипсин, окисленный фотохимическим путем, в котором разрушена имидазольная группа [77], не способен реагировать с ДФФ. [9]
Автор подробно останавливается на кинетике и термодинамике процессов ингибирования холинэстераз. Рассматривается участие гистидина и серина активных центров эстераз в процессе фосфорилирования ферментов. Особый интерес представляют обобщенные данные по изучению связи между строением и антихолинэстеразной активностью фосфорорганических ингибиторов. Приводятся данные по использованию уравнения Гаммета для нахождения зависимости между антихолинэстеразной активностью и структурой ингибиторов. Описываются фосфорорганические ингибиторы, содержащие в своем составе катионный или катионогенный участки. Представляют особый интерес данные о различной антихолинэстеразной активности оптических изомеров этилового эфира S - 3-этилмеркапто-этилтиолоэтилфосфоната. [10]
Как правило, при киназной реакции осуществляется непосредственный перенос фосфор-ильной группы от донора к акцептору. Крейн [ И ] собрал доказательства того, что у многих киназ не происходит промежуточное фосфорилирование фермента. [11]
Очевидно, токсичность фосфорорганического соединения для организма будет заметно снижаться в результате присутствия ферментов, способных метаболизовать соединение до нетоксичных продуктов. Метаболитиче-ская детоксикация осуществляется в значительной степени катализируемым щелочным гидролизом, который происходит по лабильным связям, разрываемым при фосфорилировании фермента, в результате чего образуется диалкильный эфир фосфорной кислоты и спирт или фенол. [12]

В большинстве тканей при концентрации уабаина 0 1 - 1 мкмоль / л наблюдается 50 % - ное ингибирование Na, К - АТРазы. При этом в основном ингибируется К - зависимое де-фосфорилирование ( рис. 7.8), однако при более высокой концентрации стероидов может блокироваться и Na, М § 2 -зависи-мое фосфорилирование фермента. Ингибирование происходит на внешней стороне мембраны. [14]
АТФазами), к к-рым относятся Н - АТФаза мембран митохондрий, хло-ропластов и бактериальных клеток, Са 1 - АТФаза внутриклеточных мембран мышечных клеток ( мембран саркоплаз-матич. К - АТФаза, содержащаяся практически во всех плазматич. В результате создается неравновесное распределение ионов и генерируется разность электрич. АТФаз включает связывание АТФ и ионов на поверхности фермента, фосфорилирование фермента, перенос ионов через мембрану, отщепление ортофосфата от белка, изменение прочности связи ионов с ферментом, возврат системы в исходное состояние. Процессы перемещения ионов через мембраны связаны с изменением пространств, структуры белковой части трансп. АТФаз, однако детальный механизм процесса пока не установлен. По-видимому, в состав АТФаз входят ферментный центр, ионный канал и какие-то структурные элементы, препятствующие обратной утечке ионов во время работы И. В мембранах митохондрий, хлоропластов и клеток бактерий функционирует и др. механизм активного переноса протонов: сопряжение трансмембранного переноса протонов с переносом электронов через определ. Работа таких систем вместе с работой обращенной Н - АТФазы приводит к окислительному фосфорили-рованию в митохондриях и у бактерий и к фотосинтетич. [15]
Страницы: 1