Фрагмент - полимер
Cтраница 1
Фрагмент полимера, который мы называем целлюлозой, представлен ниже. Это пример полимерного циклического ацеталя, схематически пока-ванного выше. [1]
Металлпорфириновые фрагменты полимера в значительной степени ведут себя как независимые комплексы, о чем свидетельствуют их спектральные и каталитические свойства, хотя и проявляется сильное коллективное взаимодействие в высокосопряженной системе полимера. [2]

Деструкция фрагментов полимеров с насыщенными связями под действием озона протекает через образование пероксидных радикалов. [4]

Деструкция фрагментов полимеров с ненасыщенными связями ( полидиенов) происходит через образование озо-нида и цвиттер-ионов. [6]
Анализ различий в энергии я-электронов между двойными анион-радикальными фрагментами полимера и образующимися из них анионами показывает, что постулируемая в этом механизме диссоциация связи С - С энергетически выгодна. Эти значения, вычисленные методом МО в приближении Хюккеля, приведены в табл. 59 в порядке увеличения выигрыша в энергии я-электронов. Если основным фактором, определяющим способность полимера к распаду по данной схеме, является выигрыш в энергии л-электронов, то с помощью табл. 59 легко установить шкалу относительной устойчивости полимерных анион-радикалов, которая согласуется с экспериментальными данными. [7]
Негомогенная активность одного или нескольких реагентов из-за накопления его молекул вблизи определенных фрагментов полимера чаще наблюдается в процессе полимераналогичных превращений в гомогенных системах. Этот эффект может проявляться в виде водородных или ионных связей, а также гидрофобных взаимодействий. Негомогенная активность реагентов может быть обусловлена также кооперативными эффектами. Наряду со специфическим воздействием среды, например гидрофобным воздействием и кооперативными эффектами, негомогенное распределение активности играет большую роль при гомогенном катализе на макромолекулах. [8]
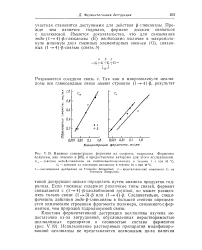 |
Влияние концентрации фермента на скорость гидролиза. Ферменты. [9] |
Следовательно, специфичность действия эндо-р-глюканазы в большей степени определяется химическим строением фрагмента полимера, связанного с ферментом, чем природой гидролизуемой связи. [10]
В случае полимеров с температурой стеклования намного выше комнатной ( например, полиметилметакрилат) даже крошечные фрагменты полимера, составляющие ядро мицеллы, могут пребывать ниже его температуры стеклования в полурастворителе и, следовательно, быть достаточно жесткими. В обычных мицеллах, хотя ассоциация якорных цепей энергетически крайне благоприятна, тем не менее энергия ассоциации не слишком велика по сравнению с kT и для разрушения мицеллы не требуется дополнительной энергии активации. [11]
Следовательно, чтобы мог произойти внутриклеточный гидролиз, необходимо структуры органического матрикса предварительно подвергнуть воздействию, в результате которого образовались бы фрагменты полимеров. Так, резорбция коллагеновых волокон требует предварительного воздействия коллагенолитических ферментов. До недавнего времени считали, что коллагеназа отсутствует в животных тканях. Рядом исследователей доказано присутствие коллагенолитических ферментов в некоторых тканях животных, в частности в костной ткани. [12]
В результате получаются иные, чем в случае III, продукты: не аде привитые ветви А, а привитая ветвь А и стабильный фрагмент полимера А. [13]
В результате получаются иные, чем в случае III, продукты: не две привитые ветви А, а привитая ветвь А и стабильный фрагмент полимера А. [14]
Перенос энергии растворитель - растворенное вещество в пластических растворах был рассмотрен аналогичным образом [15, 118], но остался не вполне решенным вопрос о том, дает ли какой-либо вклад в эффективность переноса миграция энергии между фрагментами полимера. Крайне желательно, чтобы были получены спектры поглощения и испускания отдельных компонент какой-либо исследованной сцинтилляционной системы. Это дало бы возможность провести вычисления, связанные с переносом энергии, и различить первичные и вторичные процессы сцинтилляции. [15]
Страницы: 1 2