Хеморецептор
Cтраница 1
 |
А. Система трахей у саранчи. Б. Строение трахеи насекомого.| Условия, создающиеся в тканях насекомого в покое и в активном состоянии ( работа трахеол. [1] |
Хеморецепторы улавливают это и дыхальца открываются. Тот же стимул может вызывать и вентиляционные движения тела, особенно у крупных насекомых, таких как саранча. Всасывание воздуха ( вдох) происходит пассивно, когда сегменты тела благодаря своей эластичности принимают исходную форму. [2]
 |
Условный сосудосуживающий рефлекс при сочетании звука звонка с болевым температурным раздражителем. [3] |
Хеморецепторы обнаружены также в сосудах селезенки, надпочечников, почек, костного мозга. [4]
Хеморецепторы, стимулируемые увеличением напряжения двуокиси углерода и снижением напряжения кислорода, находятся в каротидных синусах и дуге аорты. Они расположены в специальных маленьких тельцах, обильно снабжаемых артериальной кровью. Важными для регуляции дыхания являются каротид-ные хеморецепторы. Аортальные хеморецепторы на дыхание влияют слабо и имеют большее значение для регуляции кровообращения. [5]
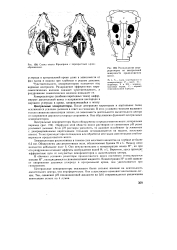 |
Расположение хемо-рецепторов на вентральной поверхности продолговатого мозга. [6] |
Хеморецепторы ( особенно каротидных телец) информируют дыхательный центр о напряжении кислорода и двуокиси углерода в крови, направляющейся к мозгу. [7]
Хеморецепторы расположены в тонком слое мозгового вещества на глубине не более 0 2 мм. Обнаружены два рецептивных поля, обозначаемые буквам М и L. Оно нечувствительно к концентрации ионов Н, но при его разрушении исчезают эффекты возбуждения полей М и L. Вероятно, здесь проходят афферентные пути от сосудистых хеморецепторов к дыхательному центру. [8]
Хеморецепторы телец особенно чувствительны к колебаниям газового состава крови. [9]
Хеморецепторы каротидных и аортальных телец являются уникальными рецептор-ными образованиями, на которые гипоксия оказывает стимулирующее влияние. [10]
Чувствительность хеморецепторов находится под нервным контролем. Раздражение эфферентных парасимпатических волокон снижает чувствительность, а раздражение симпатических волокон повышает ее. [11]
Возбуждение хеморецепторов головного мозга может быть вызвано высвобождением из нервных окончаний его структур гистамина, индольных соединений, изменением содержания в желудочках двуокиси углерода и другими факторами. Рецепторы каротид-ных клубочков реагируют на недостаток в крови кислорода, на снижение величины рН ( в пределах 6 9 - 7 6) и повышение напряжения углекислоты. [12]
Если принципы действия хеморецептора и бактериального мотора относительно ясны [9-12], то вопрос о передаче сигнала от хеморецептора к мотору ( или регулятору дрожаний) остается наименее выясненным. Повышение концентрации ионов Са вблизи мотора вызывает конформационное изменение регулятора кувырканий и приводит к изменению частоты дрожаний бактерий. [13]
В кровеносных сосудах имеются хеморецепторы, способные реагировать на появление в крови активного тромбина. Хеморецепторы связаны с ней-рогуморальным механизмом, регулирующим образование антикоагулянтов. Таким образом, если тромбин появляется в циркулирующей крови в условиях нормального нейрогуморального контроля, то в этом случае он не только не вызывает свертывания крови, но, напротив, рефлекторно стимулирует образование антикоагулянтов и тем самым выключает свертывающий механизм. [14]

Страницы: 1 2 3 4 5