Хемостат
Cтраница 2
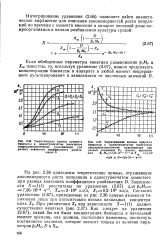
На рис. 2.36 приведены теоретические кривые, отражающие закономерности роста популяции в одноступенчатом хемостате при разных значениях коэффициента разбавления D. Как следует из уравнения (2.67), какой бы ни была концентрация биомассы в начальный момент времени, она должна, постепенно изменяясь, достигнуть постоянного значения, которое зависит только от величин параметров PlAf0, D И Хао. [17]
 |
Репрессия орнитин. карба-моил-трансферазы. Образование ферментов, необходимых для биосинтеза аргинина, при добавлении к среде аргинина почти мгновенно прекращается ( репрессируется. [18] |
На мутантах, дефектных по одному из ферментов биосинтеза аргинина, путем опытов в хемостате удалось показать, что состояние дерепрессии может сохраняться: если рост клеток лимитируется аргинином, активность фермента в 25 раз выше обычного уровня. [19]
При определении скорости роста в постоянных системах культур стерилизованная фильтрованием вода равномерно поступает из больших резервуаров в хемостат. [20]
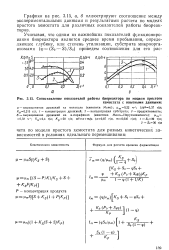 |
Сопоставление показателей работы биореактора по модели простого. [21] |
Графики на рис. 3.15, а, б иллюстрируют соотношение между экспериментальными данными и результатами расчета по моделк простого хемостата для различных показателей работы биореакторов. [22]
Типичная зависимость х и с от D для одностадийного работающего в режиме полного смешения проточного биореактора непрерывного действия или хемостата показана рис. 3.4. Из рисунка видно, что величины х и с остаются практически постоянными в широком интервале скоростей разбавления. Однако, когда D приближается к цт, то происходит крутое падение значения х с одновременным резким возрастанием с. Точка, в которой х О и с CQ, является критической скоростью разбавления, равной максимуму удельной скорости роста; после нее начинается полное вымывание биомассы в системе, работающей в режиме полного смешения. [23]
Анализ литературных данных показывает, что наиболее успешно и относительно быстро микроорганизмы адаптируются к пестицидам в перфузионных системах, хемостате, почвенной суспензии, на затопляемых почвах, а также в случаях, когда концентрация химических веществ в десятки и сотни раз превышает концентрацию их в производственных условиях. [24]
На основе установленной зависимости удельной скорости роста от концентрации субстрата в питательной среде, обратившись к проблеме непрерывного культивирования, Моно [82] обосновал принцип хемостата, послуживший теоретической основой различных приемов и способов культивирования широкого круга микроорганизмов. [25]

На рис. 5.5 показаны две высокотехнологичные системы такого рода. В одноступенчатой системе в хемостат с культурой сульфат-редуцирующих бактерий непрерывно поступает раствор, загрязненный каким-либо металлом и содержащий сульфат и питательные вещества типа лактата или в зависимости от природы используемой бактерии углеводород либо органическую кислоту. [27]
Мы видели, что как среди растений, так и среди животных существуют весьма разнообразные взаимоотношения, которые можно считать мутуалистическим симбиозом. Далее по уровню сложности следуют экосистемы типа хемостата ( строго внешние по отношению к тканям) в рубце жвачных и слепой кишке термитов; затем - межклеточная эктомикориза и внутриклеточные зооксантеллы кишечнополостных. [28]
Адаптация происходит, как правило, очень медленно. Поэтому для выделения нужных микроорганизмов из почвы обычно используют технику накопительной культуры, перколяторы и хемостаты, причем период предварительного культивирования часто растягивается на многие недели и месяцы. [29]
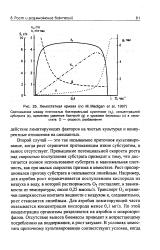 |
Хемостатная кривая ( по М. Madigan et al, 1997. [30] |
Страницы: 1 2 3