Хлоропласт - высшее растение
Cтраница 2
Исследования 30 - 40 - х годов по этому вопросу Местра ( 1930), Штоля ( 1936), Моммартса ( 1938), Штоля и Видемана ( 1938, 1941), Катца и Вассинка ( 1939), Френча ( 1942), Годнева и Оси-повои ( 1947, 1948) и ряда других ученых99 также показали действительное существование хлорофилл-белковых соединений не только в хлоропластах высших растений, но и у водорослей и бактерий. Многочисленные исследования позволили установить, что комплексные соединения простетических групп со своими белковыми носителями присущи всем растительным пигментам. [16]
Однако хлоропласта и митохондрии эукариотических клеток содержат рибосомы, отличные от 80S типа. Рибосомы хлоропластов высших растений принадлежат к истинному 70S типу и практически не отличимы от рибосом эубактерий и синезеленых водорослей по вышеприведенным показателям и по более детальным молекулярным характеристикам. Митохондриальные рибосомы более разнообразны в зависимости от принадлежности организма к тому или иному царству эукариот. Наиболее изучены рибосомы митохондрий грибов и млекопитающих. Митохондриальные рибосомы грибов ( Saccharomyces, Neurospora) похожи на прокариотические 70S рибосомы, но, может быть, лишь слегка крупнее ( около 75S) и содержат относительно больше белка; абсолютное содержание рибосомной РНК в них, по-видимому, почти такое же, как в типичных 70S рибосомах. Митохондриальные рибосомы млекопитающих, однако, существенно мельче типичных 70S рибосом, имея также и существенно меньшее абсолютное количество рибосомной РНК на частицу; их иногда называют мини-рибосомами. Действительно, коэффициент седиментации рибосом из митохондрий млекопитающих составляет всего около 55S, а тотальная масса рибосомной РНК на частицу более чем на 1 / 3 меньше, чем в типичных 70S рибосомах. В то же время, Митохондриальные рибосомы млекопитающих содержат довольно много белка, так что общие размеры их как будто бы не сильно отличаются от таковых прокариотических рибосом. В целом, несмотря на ряд необычных черт, по ряду своих признаков, и в том числе по функциональному поведению, Митохондриальные рибосомы млекопитающих все же близки к прокариотическим 70S рибосомам. [17]
В последние годы открыт еще один физиологический иереносчик электронов в фотосинтетической транспортной цепи - пластоцианин. Обнаружен в хлоропластах высших растений, г; водорослях. Не найден у фото-синтезирующих бактерий. [18]
 |
Схема строения митохондрии. [19] |
В связи с развитием электронной микроскопии в последние годы сделаны крупные успехи в изучении строения хлоропла-стов. Было показано, в частности, что хлоропласты высших растений имеют пластинчатую структуру. Пластинчатая структура наблюдается только у пигментированных хлоропластов. [20]
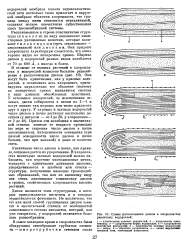 |
Схемы расположения дисков в хлоропластах различных водорослей. [21] |
Увеличение числа дисков в пачке, как правило, сопровождается их укорочением. В хлоропла-стах некоторых зеленых водорослей можно наблюдать, как короткие многодисковые пачки, чередуясь с единичными длинными дисками, упорядочиваются в штабеля или стопки - структуры, получившие название граноподоб-ных образований, так как по внешпему виду они очень напоминают компоновку дисков в гранах, свойственных хлоропластам высших растений. [22]
В каждой клетке высших растений имеется от 20 до 100 хлоропластов. У некоторых водорослей, например у хлореллы, в клетке имеется только один хлоропласт. Хлоропласты высших растений имеют эллипсоидальную форму с размерами осей от 1 до 10 мкм. У хлоропластов низших растений их форма весьма разнообразна: спиральная, звездчатая, пластинчатая. [23]
Как Прокариотическая, так и эукариотическая рибосомы содержат две различные высокополимерные РНК, по одной на каждую субчастицу, и одну относительно низкомолекулярную РНК, так называемую 58 РНК. Кроме того, эукариотические рибосомы содержат и другую относительно низкомолекулярную РНК, так называемую 5 88 РНК, которая является гомологом 5 -концевой части ( около 160 нуклеотид-ных остатков) высокополимерной РНК большой субчастицы прокариот. В рибосомах хлоропластов высших растений имеется также так называемая 4 58 РНК, которая является гомологом З - концевой части ( около 100 нуклеотидных остатков) высокополимерной РНК большой субчастицы бактерий. Таким образом, 5 88 РНК эукариоти-ческих рибосом и 4 58 РНК хлоропластных рибосом являются результатом расщепления ( процессинга) предшественника высокополимерной РНК большой субчастицы в процессе биогенеза или созревания рибосом; они непосредственно участвуют в формировании структуры высокополимерной РНК большой субчастицы, как и их гомологичные последовательности у бактерий ( см. ниже), и поэтому могут не рассматриваться как самостоятельные виды рибосомной РНК. [24]
В опытах Уиттингема и Бишопа [323] по исследованию реакции Хилла в хлоропластах шпината при 4 С не наблюдалось существенного влияния порядка облучения на выход кислорода. Одиночная вспышка монохроматического света длительностью 35 мс давалась через разные промежутки времени после выключения монохроматического света другой длины волны. Усиление, наблюдавшееся, когда вспышка 697 нм давалась после некоторого периода облучения светом 653 нм, было лишь немногим меньше усиления, наблюдавшегося, когда вспышка 644 нм следовала за периодом облучения светом 700 нм ( фиг. Кроме того, мы сравниваем здесь красную водоросль, содержащую фикоэритрин и не содержащую хлорофилла Ъ, с хлоропластами высшего растения, в которых имеется хлорофилл Ь, но нет фи-кобилина. Первое из упомянутых отличий может иметь решающее значение, и любопытные результаты Френча, возможно, объясняются тем, что зеленый продукт окисляется вследствие стимуляции дыхания более коротковолновым светом. [25]
В организме они синтезируются димеризацией геранилгеранилпирофосфата по схеме хвост к хвосту. Они подразделяются на каротины - собственно углеводороды - и ксантофиллы - гидрокси -, метокси -, эпокси -, оксо - и другие кислородсодержащие производные каротинов. Биосинтез ксантофиллов идет с участием кислорода воздуха, а не воды. При этом углеводородная цепь их может быть циклизована; различают моноциклические ( с одним замкнутым шестичленным кольцом) и би-циклические каротиноиды. Если в хлоропластах высших растений присутствуют только бициклические формы каротиноидов, то у зеленых растений и водорослей - моно - и бициклические, а у бактерий - только ациклические формы. [26]
Страницы: 1 2