Центросома
Cтраница 2
Что касается сложных явлений, происходящих в ядре клетки при ее не прямом кариокинетическом делении, то они весьма тщательно описаны гистологами, открывшими существование особых областей, занятых форменными элементами клетки - центросомами. При изучении морфологии кариокинеза создается впечатление, что центросомы представляют собой образования, притягивающие части делящегося ядра. Попытки как морфологов, так и биофизиков открыть природу сил, возникающих между центросомами и хроматином ядра, не дали до сих пор удовлетворительных результатов, и физико-химическое объяснение процессов деления клетки, лежащего в основе огромного числа биологических явлений, является до сих пор недостигнутым. [16]

Центриоли всегда бывают расположены в материале, не имеющем четко выраженной структуры, который инициирует развитие микротрубочек. Эту область клетки называют центросомой. Именно центросома образует веретено, потому что нити веретена на самом деле представляют собой микротрубочки. Это позволяет объяснить, как растения и грибы, не имеющие центриолей, также способны образовывать веретено. Функция центриолей в делении ядра остается неясной. Возможно, они участвуют в ориентации веретена, помогая таким образом установить, в какой плоскости будет проходить деление клетки. Некоторые нити веретена тянутся от одного полюса к другому, тогда как другие - от полюсов к центромерам. Укорочение этих нитей веретена в результате удаления субъединиц тубулина позволяет объяснить перемещения хромосом и хроматид во время клеточного деления. Они фактически наматываются центросомами. [18]
Что касается сложных явлений, происходящих в ядре клетки при ее не прямом кариокинетическом делении, то они весьма тщательно описаны гистологами, открывшими существование особых областей, занятых форменными элементами клетки - центросомами. При изучении морфологии кариокинеза создается впечатление, что центросомы представляют собой образования, притягивающие части делящегося ядра. Попытки как морфологов, так и биофизиков открыть природу сил, возникающих между центросомами и хроматином ядра, не дали до сих пор удовлетворительных результатов, и физико-химическое объяснение процессов деления клетки, лежащего в основе огромного числа биологических явлений, является до сих пор недостигнутым. [19]
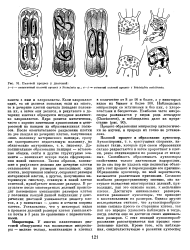 |
Половой процесс у диатомей. [20] |
Если хлоропласт один, то он делится пополам; если их много, то в дочерние клетки сначала попадает половина их, а затем они делятся, в результате в дочерних клетках образуется исходное количество хлоропластов. Ядро делится митотически, часто с хорошо заметными хромосомами и центросомой на каждом из образовавшихся полюсов. Дополнительные образования панциря - вставочные ободки, септы и другие структурные элементы - возникают вскоре после сформирования новой гипотеки. В результате после многократных делений происходит постепенное уменьшение размеров клеток у половины каждой данной популяции: у центрических диатомей уменьшается диаметр клеток, а у пеннатных - длина и отчасти ширина клеток. Установлено, что у некоторых видов в процессе делений размеры клеток уменьшаются почти в 3 раза по сравнению с первоначальными. [21]
Другой органеллой, в состав которой входят гладкие мембраны и которая функционально, а возможно, и структурно связана с эндоплазматической сетью, является аппарат, или комплекс Гольджи. Он обычно расположен совсем близко от ядра ( в области так называемых центросом, или центросфер) и состоит из уложенных в стопку гладких мешочков, а также из различного числа цистерн, пузырьков и вакуолей с гладкой поверхностью. Хорошо сформированные комплексы Гольджи в большом количестве встречаются в секреторных клетках, таких, как экзокринные клетки поджелудочной железы. Убедительные данные указывают на существование связи между цистернами гранулярной сети и пузырьками комплекса Гольджи, которые в свою очередь связаны с более крупными вакуолями комплекса. Вакуоли Гольджи дают начало секреторным гранулам, например зимогеновым гранулам, в которых содержатся и накапливаются белки, синтезированные рибосомами гранулярной сети ( фиг. [22]
Центриоли всегда бывают расположены в материале, не имеющем четко выраженной структуры, который инициирует развитие микротрубочек. Эту область клетки называют центросомой. Именно центросома образует веретено, потому что нити веретена на самом деле представляют собой микротрубочки. Это позволяет объяснить, как растения и грибы, не имеющие центриолей, также способны образовывать веретено. Функция центриолей в делении ядра остается неясной. Возможно, они участвуют в ориентации веретена, помогая таким образом установить, в какой плоскости будет проходить деление клетки. Некоторые нити веретена тянутся от одного полюса к другому, тогда как другие - от полюсов к центромерам. Укорочение этих нитей веретена в результате удаления субъединиц тубулина позволяет объяснить перемещения хромосом и хроматид во время клеточного деления. Они фактически наматываются центросомами. [23]
У большинства животных и у некоторых низших растений в образовании веретена принимает участие специальное тельце, так называемая центросома, которая во время стадии покоя и профазы лежит вне ядерной оболочки. Таким образом, они, по-видимому, имеют важное значение для развития веретена. У цветковых растений центросом нет, однако веретено образуется у них столь же регулярно, как и у видов, имеющих центросому. [24]
Основные стадии митоза изображены на фиг. В профазе молекулы ДНК ( ген) реплицируются, хромосома утолщается и удваивается; при этом хромосомы отчетливо вырисовываются в виде двойных нитевидных тел, удерживаемых вместе центромерами. В поздней профазе ядерная оболочка разрушается, а между центросомами образуется веретено. В метафазе неразделившиеся центромеры все еще удерживают вместе хромосомы, однако теперь хромосомы расположены по экватору веретена. В анафазе центромеры разделяются и отходят к противоположным полюсам, увлекая за собой и дочерние хромосомы. В телофазе начинается деление клетки. Намечается образование клеточных мембран. После телофазы наступает новая интерфаза - образуются две дочерние клетки с полным набором хромосом ( и генов), которые идентичны между собой и с хромосомами ( и генами) родительской клетки. В клетке способны к репликации, по-видимому, только хромосомы. Поскольку в репликации зиготы участвуют гены обоих родителей, становится понятным, каким образом достигается строгая передача единиц наследственности от одного поколения другому. [25]
В предыдущих главах мы объяснили роль хромосом в наследственности и нашли, что все развитие генетики тесно связано с исследованием свойств и эффектов генов и хромосом. Хотя в этой области достигнуты большие результаты, нельзя забывать, что существуют и другие клеточные компоненты, которые могут играть важную роль в наследственности. Кроме ядра, в клетке имеются другие элементы, размножающиеся делением, например центросомы, которые у животных и у низших растений участвуют в образовании ядерного веретена ( см. стр. Как хорошо известно, хлорофилльные зерна осуществляют важнейший процесс - ассимиляцию углекислоты, благодаря которому становится возможным синтез сахара и крахмала из воды и углекислоты воздуха. Пластиды способны к самовоспроизведению, подобно хондриосомам, которые встречаются как в клетках растений, так и в клетках животных. [26]
Что касается сложных явлений, происходящих в ядре клетки при ее не прямом кариокинетическом делении, то они весьма тщательно описаны гистологами, открывшими существование особых областей, занятых форменными элементами клетки - центросомами. При изучении морфологии кариокинеза создается впечатление, что центросомы представляют собой образования, притягивающие части делящегося ядра. Попытки как морфологов, так и биофизиков открыть природу сил, возникающих между центросомами и хроматином ядра, не дали до сих пор удовлетворительных результатов, и физико-химическое объяснение процессов деления клетки, лежащего в основе огромного числа биологических явлений, является до сих пор недостигнутым. [27]
У большинства животных и у некоторых низших растений в образовании веретена принимает участие специальное тельце, так называемая центросома, которая во время стадии покоя и профазы лежит вне ядерной оболочки. Таким образом, они, по-видимому, имеют важное значение для развития веретена. У цветковых растений центросом нет, однако веретено образуется у них столь же регулярно, как и у видов, имеющих центросому. [28]
В процессе митотического деления хорошо различимы несколько фаз. В течение первой фазы, или профазы, в ядре появляются нитевидные структуры, хроматиды, а ядрышко исчезает. Эти хроматиды в конечном итоге путем сокращения и скручивания формируют интенсивно базофильные компактные хромосомы. В течение этой фазы центросомы, расположенные вне ядра, делятся на две половины, которые расходятся к противоположным сторонам ядра. Ядерная мембрана и ядрышко исчезают, возникает структура, называемая веретеном. Она состоит из тонких нитей, расходящихся от каждой центросомы к экватору веретенообразной фигуры. В течение метафазы хромосомы выстраиваются у экватора веретена, и каждая из них делится на две равные части, которые на протяжении анафазы расходятся к противоположным полюсам веретена. В результате этого процесса каждая дочерняя клетка получает тот хромосомный материал, который имелся у материнской клетки. В течение последней фазы деления - телофазы - образуются новые ядра. При этом из каждой группы дочерних хромосом, теряющих свои очертания, формируется хроматин нового ядра; одновременно образуются новая ядерная мембрана и ядрышко. [29]
Участок лучистой фигуры, лежащей между центросомами, постепенно удлиняясь, приобретает форму веретена. К этому времени ядерная оболочка исчезает, пропадает ядрышко, а ядерный сок смешивается с протоплазмой. Теперь вместо круглого ядра в центре клетки мы видим только хромосомы, расположенные в средней части лучистого веретена. Оно образовано тонкими нитями, как бы натянутыми между центросомами. Дальше наступает один из наиболее существенных и замечательных моментов деления ядра. Он заключается в том, что хромосомы расщепляются вдоль на две половины, то есть из каждой материнской хромосомы образуются две абсолютно сходных дочерних. Общее число хромосом, таким образом, удваивается: если их было четыре, то в результате деления получается восемь. [30]
Страницы: 1 2 3